Caractéristiques générales des annélides. Vers annelés : photo, description et mode de vie
Annélides - animaux multicellulaires, à symétrie binaire, à trois couches qui ont une cavité corporelle secondaire. Le type Kilchakiv regroupe plus de 9 000 espèces (en Ukraine - environ 450 espèces). Ils vivent dans la mer et les eaux douces, ainsi que dans le sol, par rapport aux représentants d'autres types de vers, les kilchakids ont un niveau d'organisation nettement plus élevé. Les caractéristiques progressives de l'organisation des annélides sont : 1 ) apparition d'une cavité corporelle secondaire 2) division du corps en segments séparés (métamérisme) ; 3 ) apparition de membres primitifs (parapodes chez les vers polychètes) ; 4 ) aspect des systèmes circulatoire et respiratoire (branchies externes chez les vers polychètes) ; 5 ) développement de métanéphridie.
caractéristiques structurelles
Corps en kilchak multicellulaire, formé de plusieurs segments. La répétition répétée des segments du corps est appelée métamérisme. Ce principe d'organisation du corps apparaît dans le processus d'évolution en lien avec l'allongement du corps. Segments - parties du corps de structure similaire, situées séquentiellement les unes après les autres. Les segments à l'extérieur et à l'intérieur sont les mêmes, donc à Kilchakiv Le métamérisme, ou segmentation, du corps est homonomique. De nombreux représentants ont des soies sur les segments du corps. Le corps est constitué de la tête, du tronc et du lobe anal. Une division aussi claire du corps en sections est observée pour la première fois à Kilchakiv.
Voiles sont représentés par un sac cutanéo-musculaire bien développé, qui comprend : dense, fin cuticule , hypoderme Et deux couches de muscles(circulaire et longitudinal). Il existe de nombreuses glandes muqueuses dans la peau.
Cavité corporelle secondaire (en général) segmenté. Elle se distingue de la cavité primaire par la présence de son propre revêtement épithélial, adjacent aux parois du sac d'un côté et aux parois du tube digestif de l'autre. Les feuilles qui tapissent se développent ensemble au-dessus et au-dessous du tube, formant un mésentère qui divise l'ensemble en parties droite et gauche. Des cloisons transversales divisent la cavité corporelle en chambres dont le nombre correspond au nombre de segments. L’ensemble est rempli d’un liquide dont la composition chimique est très proche de l’eau de mer. Le liquide coelomique est en mouvement constant et remplit un certain nombre de fonctions importantes : laver les organes du corps, leur fournir des nutriments et de l'oxygène avec le sang, favoriser l'élimination du CO2 et des produits métaboliques, déplacer les substances biologiquement actives et les phagocytes, etc.
caractéristiques de la vie
Soutien associé à hydrosquelette, fonctionnant grâce au fluide cœlomique.
Mouvement musculaire. Chez les vers polychètes, il existe des segments du corps en croissance musculaire - parapodes, qui forment l’appareil moteur. Chez les oligochètes, des touffes de poils les remplacent.
Digestion est assuré par un système digestif différencié, qui comporte trois sections : antérieure (bouche, pharynx, œsophage avec une hache, estomac avec des parois musculaires), moyenne (intestin moyen) et postérieure (intestin postérieur avec anus). Chaque département du système remplit sa propre fonction spéciale. Par exemple, dans la paroi de l'intestin moyen se trouvent des cellules qui sécrètent des enzymes digestives et des cellules qui digèrent les aliments. La fonction principale de cette section est donc la digestion et l'absorption des aliments.
Transport de substances à Kilchakiv est déjà en cours avec la participation système circulatoire, qui apparaît pour la première fois. Chez les annélides système circulatoire fermé - un système dans lequel le sang circule uniquement dans les vaisseaux et ne pénètre pas dans la cavité corporelle. Leur système circulatoire est formé de vaisseaux sanguins dorsaux et abdominaux reliés entre eux par des vaisseaux annulaires. De ces vaisseaux partent de petits capillaires qui, en se ramifiant, forment un réseau dense dans la peau et les organes internes. Le mouvement du sang est dû à la pulsation des vaisseaux annulaires ; ils n'ont pas de cœur. Le sang peut être incolore ou coloré en raison de la présence de pigments respiratoires : chlorocruorine (détermine la couleur verte du sang), hémoglobine et hémoérythrine (détermine la couleur rouge).
Haleine est déjà réalisé avec la participation système respiratoire, qui apparaît pour la première fois. son apparition est associée à un métabolisme intensif par rapport aux groupes d'invertébrés précédents. Dans certains coquillages, des organes respiratoires aquatiques apparaissent - branchies, constituant des formations à parois minces avec un vaste réseau de vaisseaux, situés sur les parapodes, la tête et la queue. Mais dans la plupart des kilchaks, les échanges gazeux se font à travers le tégument.
Sélection se produit avec la participation du système excréteur formé par des organes spéciaux - métanéphridie. Ces organes commencent dans la cavité corporelle sous la forme d’un entonnoir à partir duquel s’étend un canal s’ouvrant vers l’extérieur dans un autre segment. Chaque segment contient une paire de ces organes excréteurs.
Règlement processus réalisée par un système nodal nerveux de type chaîne. Le système nerveux central se forme ganglions nerveux suprapharyngés et sous-pharyngés, jonctions navcolopharyngées et cordon nerveux ventral. Le PNS est représenté par des branches nerveuses.
Irritabilité fournir des organes sensoriels bien développés. Dans le tégument se trouvent des cellules sensibles qui distinguent les goûts et les odeurs ; les organes du toucher sont les antennes, les soies, les organes existants de la vision et parfois les organes de l'équilibre.
Reproduction principalement sexuelle avec la participation du système reproducteur. Les vers polychètes sont dioïques, tandis que les oligochètes et les sangsues sont hermaphrodites. La fécondation peut être externe ou interne. La reproduction asexuée se retrouve également chez les annélides aquatiques, dont le corps peut se fragmenter en plusieurs parties inégales (division désordonnée) ou en segments séparés (fragmentation multiple).
Développement chez les oligochètes et les sangsues - droits. Chez les polychètes - indirect, dans lequel se forme une larve trochophores. Il flotte dans l'eau pendant un certain temps, puis se dépose au fond et se transforme en un organisme adulte.
Régénération bien développé chez les oligochètes et les polychètes, mais chez les sangsues cette capacité est perdue.
Variété d'annélides
Le phylum est divisé en plusieurs classes, dont les polychètes, les oligochètes et les sangsues.
Vers polychètes, ou polychètes - une classe d'annélides qui ont des parapodes avec de nombreuses soies sur chaque segment du corps. Cette classe comprend environ 5 300 espèces qui vivent principalement dans les mers, et seuls quelques représentants se sont adaptés à la vie dans les plans d'eau douce ou les zones humides. Ils mènent une vie de fond (animaux benthiques), certains nagent librement, il existe également des formes sessiles et sécrètent des tubes protecteurs autour d'eux. Principalement des prédateurs, mais parmi eux il existe de nombreuses formes herbivores et omnivores. Parmi les polychètes, il existe également des espèces commensales qui vivent à l'intérieur des éponges, dans la carapace des bernard-l'ermite ou sur des étoiles de mer. Le corps des vers polychètes se compose d'une section de tête, d'un corps segmenté et d'un lobe anal. Sur la tête se trouvent des yeux, des organes du toucher - des tentacules, des organes de l'odorat - des fosses odorantes et, dans certains cas, des organes d'équilibre (statocystes). Sur les segments du corps se trouvent des excroissances latérales appariées avec des poils - des parapodes - des membres primitifs à l'aide desquels les vers nagent, rampent ou s'enfouissent dans le sol. Les chaètes riches ont principalement une respiration cutanée, mais certaines espèces ont des branchies. La plupart des vers polychètes sont des animaux dioïques. La fécondation des œufs se produit dans l'eau. Le développement est indirect, dans lequel des larves de trochophores libres émergent des œufs, poussées par le mouvement des cils. Les représentants les plus célèbres de la classe sont la veine de sable, la néréide et le palolo.
Grès marin (Arenico/o marina) - un ver polychète marin qui vit dans la mer Noire et peut atteindre 30 cm de long. Il passe toute sa vie dans un trou profond fait de sable. Il se nourrit de petites algues, d'animaux et de divers morceaux inanimés, les capturant avec du sable. C'est l'aliment principal du poisson commercial.
Néréis (Néréis virens) , ou Néréide,- ver polychète vivant dans la mer d'Azov. Fait référence au benthos - rampe le long du fond marin à l'aide des paramètres d'événement. C'est l'aliment principal du poisson commercial. Acclimaté dans la mer Caspienne, il s'y multiplie intensément et devient un élément important du régime alimentaire des esturgeons.
Palolo (Eunice viridis) - un ver polychète marin qui vit dans les récifs coralliens des îles tropicales de l'océan Pacifique. La couleur du corps est palolo verdâtre, la taille peut atteindre 1 m. Deux fois par an, à une certaine phase de la Lune, des individus sexuellement matures apparaissent en grand nombre à la surface de l'eau pour se reproduire.
Oligochètes ou oligochètes- un groupe d'annélides qui possèdent quelques soies sur chaque segment du corps. Cette classe comprend plus de 5 000 espèces, qui habitent principalement les eaux douces et le sol, et sont beaucoup moins communes en eau salée. La plupart des oligochètes ont des tailles allant de 0,5 mm à 40 cm, et certains types de vers de terre tropicaux atteignent 3 m. Les caractéristiques de la structure externe des oligochètes sont la segmentation du corps (de 5 à 6 à 600 anneaux), l'absence de parapodes (à leur place). sont des bouquets de quelques soies), la présence d'une ceinture glandulaire dans la partie antérieure du corps chez les individus sexuellement matures, etc. Les représentants les plus connus des oligochètes sont les vers de terre et les vers tubulaires.
Ver de terre commun (Lumbricus terrestre) - une espèce de vers oligochètes qui s'est adaptée à la vie dans le sol. Ces vers appartiennent à la famille des vrais vers de terre, qui comprend environ 300 espèces. La taille du corps varie de 2 à 50 cm. Dans le sud de l'Ukraine, les vers atteignent de grandes tailles. Le mouvement dans le sol est facilité par un corps allongé et segmenté, pointu aux deux extrémités. Ils se déplacent en contractant et en relâchant alternativement les muscles circulaires et longitudinaux du sac musculo-cutané. Les poils dirigés vers l'arrière (8 sur chaque segment) permettent de s'accrocher aux moindres irrégularités du sol. Le mucus sécrété par les glandes cutanées réduit la friction du corps du ver, l'empêche de se dessécher, favorise la respiration et a
Structure interne d'un ver de terre : A - cuticule; B - hypoderme; B - muscles circulaires ; G - muscles longitudinaux; D - sac cutanéo-musculaire ; E - épithélium tapissant; Oui - chaîne nerveuse ventrale ; F - vaisseau sanguin abdominal ; AVEC - poils; ET - métonéphridie; ET - paroi intestinale ; À - en général; L - vaisseau sanguin dorsal
propriétés antibiotiques. Les vers de terre se nourrissent de débris végétaux morts contenus dans le sol. Dans le sol, les vers de terre creusent des tunnels profonds jusqu'à 2 m de la surface de la terre. Par temps chaud et humide, ils rampent hors de leurs terriers la nuit, recherchent des feuilles mortes humides, des brins d'herbe à moitié pourris et tirent tout cela dans leurs terriers. Les vers de terre sont donc des saprophages typiques. Ils avalent également de la terre, visible dans leurs intestins. Les vers rejettent le sol traité à la surface sous forme de tas caractéristiques dont l'apparition au printemps indique le début de l'activité des vers. On calcule qu'en un jour chaque ver fait passer dans ses intestins une quantité de terre égale au poids de son corps. Plus de 50 espèces de vers de terre sont connues en Ukraine, certaines d'entre elles sont répertoriées dans le Livre rouge de l'Ukraine (par exemple, Eizenia Gordeeva).
Pipetier ordinaire (tubifex tubifex) - ver mince rouge d'eau douce mesurant 2 à 5 cm. Vit dans les sols boueux des plans d'eau frais, très pollués et pauvres en oxygène. L'extrémité avant du ver est immergée dans le limon et l'extrémité arrière sort et se tortille continuellement - ces mouvements fournissent un afflux d'eau douce nécessaire à la respiration. Ils peuvent résister à une pollution importante des plans d’eau par diverses substances (produits pétroliers, pesticides…). La peau du dos du corps possède de nombreux capillaires sanguins. Un tube se forme autour de cette partie du corps, constitué de particules de limon collées avec le mucus du ver. Avale le sable et le limon et absorbe les nutriments qu'il contient. Il est utilisé comme aliment pour les poissons d'aquarium et constitue une source nutritionnelle pour les animaux d'eau douce, en particulier les poissons.
Sangsue médicale (Hirudo médicinal) a un corps segmenté de 8 à 12 cm de long. Sur la face dorsale sombre du corps, il y a un motif caractéristique de trois paires de rayures longitudinales rouge rouille ou rouge-jaune. Vit dans de petits réservoirs debout au fond boueux, envahis par la végétation. Il se nourrit du sang des amphibiens et des mammifères. La ventouse arrière est utilisée pour la fixation, tandis que la ventouse avant, dotée de mâchoires et de dents, est utilisée pour l'alimentation. Dans la cavité buccale, située au bas de la ventouse antérieure, se trouve trois mâchoires. Ils coupent la peau de l'animal dans laquelle la sangsue s'est attachée. Le matin, de la salive contenant de l'hirudine est libérée. Hirudine- une substance produite par les glandes salivaires de la sangsue pour empêcher la coagulation du sang. Le sang traité avec la salive de sangsue peut être stocké longtemps en réserve dans les grandes poches de ses intestins - grâce à cela, l'animal peut mourir de faim pendant longtemps (de plusieurs mois à un an). La sangsue médicinale est un hermaphrodite caractérisé par un développement direct. Elle pond ses œufs en grappes près de l'eau (mais pas dans l'eau), dans des endroits sombres et humides. Utilisé en médecine pour les maladies du système circulatoire, la gangrène et la transplantation d'organes.

Le corps des boucles est divisé en la section de la tête ( prostomium), les sonneries suivantes (ou segments, ou métamères), dont le nombre est généralement important (plusieurs dizaines), et la partie postérieure (lobe anal, ou pygidium). La section de la tête des vers marins, appelés polychètes, est bien définie et porte divers appendices : larges, étroits, etc. (Fig. 61). Dans les boucles d'eau douce et terrestres, la section de la tête est faiblement exprimée (Fig. 61). Plusieurs anneaux antérieurs peuvent être fusionnés avec le prostomium. Les segments du corps ont généralement une structure similaire. Ce type de division s'appelle segmentation homonomique ou métamérisme d'homonomie. Il est non seulement externe, mais profondément interne, puisque chaque segment est séparé des segments voisins par des cloisons et possède un ensemble d'organes.
La peau est constituée d'un épithélium monocouche et d'une fine cuticule sécrétée par celui-ci (Fig. 62). Il existe de nombreuses glandes dans la peau qui sécrètent du mucus, qui facilite le mouvement des vers, et d'autres sécrétions (par exemple, les substances qui aident à attirer les femelles vers les mâles des teignes dioïques, sont toxiques pour d'autres animaux, etc.).
Système nerveux. Ce système est bien mieux développé que celui des autres vers, et sa structure reflète très clairement la division du corps de la boucle en segments. Sa section centrale est constituée, en règle générale, de deux nœuds céphaliques situés sur la face dorsale, des cordons péripharyngés, qui se transforment sur la face ventrale en une chaîne, généralement très longue et formant un nœud dans chaque segment (Fig. 63, B), ce qui explique son nom. Ainsi, la chaîne abdominale était formée de deux brins. Dans les formes inférieures du type, les cordons restent séparés sur toute leur longueur et sont reliés par des ponts qui ressemblent à une échelle (Fig. 63, A). Un tel système est moins centralisé, il est similaire au système nerveux central des vers inférieurs - plat et primitif (voir Fig. 31, B et 54).
Les nœuds et cordons des annélides typiques sont bien mieux développés et leur structure est plus complexe que celles de ces dernières. L'ensemble du système central des boucles est séparé de l'épiderme, tandis que chez les vers inférieurs, il est toujours connecté à l'épiderme. Chaque nœud de la chaîne abdominale innerve et affecte le fonctionnement des organes situés dans l'anneau où se trouve le nœud. Les nœuds de tête, mieux développés que les nœuds de la chaîne, coordonnent le travail de ces derniers et, à travers eux, l'activité de tout le corps. De plus, ils innervent les yeux et d’autres organes sensoriels situés dans la tête du corps.
Les sens sont variés. Les cellules tactiles sont dispersées dans la peau, particulièrement nombreuses sur les appendices du corps. Il existe des organes qui perçoivent les irritations chimiques. Toutes les annélides ont des organes sensibles à la lumière. Les plus simples d'entre eux sont représentés par des cellules spéciales dispersées dans toute la peau. Par conséquent, presque toutes les teignes ont une peau sensible à la stimulation lumineuse. À l'extrémité antérieure du corps, et chez un certain nombre de sangsues à l'extrémité postérieure, les organes sensibles à la lumière deviennent plus complexes et se transforment en yeux. Un certain nombre de formes ont des organes d'équilibre dont la structure est similaire à celle des organes similaires des méduses et d'autres animaux inférieurs.
Le développement progressif du système nerveux des annélides assure des mouvements plus complexes et énergiques de leur corps, un travail actif de tous les systèmes organiques, une meilleure coordination des fonctions de toutes les parties du corps, un comportement plus complexe et permet une adaptation plus subtile de ceux-ci. animaux dans l'environnement.
Système de propulsion. Ce système chez les annélides est plus avancé que chez les vers précédemment étudiés. Le mouvement ciliaire n'est caractéristique que des larves ; chez les formes adultes, à de rares exceptions près, il est absent et leur mouvement s'effectue uniquement grâce au travail des muscles. Le sac cutanéo-musculaire est bien mieux développé que chez les vers plats et les vers protocavitaires (cf. Fig. 32, 53 et 62). Sous l'épiderme se trouve une couche bien développée de muscles circulaires (Fig. 62), constituée de longues fibres dotées de noyaux. Lorsque ces muscles se contractent, le corps du ver devient plus fin et plus long. Derrière les muscles circulaires se trouve une couche beaucoup plus épaisse de muscles longitudinaux, dont la contraction raccourcit le corps et le rend plus épais. La contraction unilatérale des muscles longitudinaux et de certains autres muscles entraîne une flexion du corps et un changement de direction du mouvement. De plus, il existe des muscles allant de la face dorsale à la face abdominale : muscles passant par les septa séparant les anneaux ; muscles de divers appendices du corps, qui jouent un rôle de soutien dans le mouvement des vers, etc. La force des muscles du sac cutanéo-musculaire est grande et permet aux vers de pénétrer rapidement en profondeur dans le sol. De nombreuses annélides savent nager. Le support des muscles est principalement constitué par l'hydrosquelette formé par le liquide de la cavité corporelle, ainsi que par les formations frontalières.
Le mouvement des annélides est facilité par des appendices auxiliaires (voir Fig. 61, 62, 64) : poils(disponible dans la grande majorité des espèces) et parapodes(disponible dans la plupart des vers marins). Les poils (voir Fig. 62, 64, A, B) sont des formations solides de matière organique, un glucide très complexe - la chitine, de différentes formes, épaisseurs et longueurs. Les poils sont formés et entraînés par des faisceaux musculaires spéciaux. Les soies sont disposées (individuellement ou en touffes) en rangées longitudinales régulières sur presque tous les anneaux des vers. Les parapodes (Fig. 64, B) sont de puissantes excroissances latérales du corps dotées de muscles bien développés. Les parapodes sont reliés de manière mobile au corps et ces appendices agissent comme un simple levier. Chaque parapode est généralement constitué de deux lobes : dorsal et ventral, qui, à leur tour, peuvent être divisés en lobes de second ordre. À l’intérieur de chacune des lames principales se trouve un poil de support. Les parapodes portent des touffes de poils qui s'étendent bien au-delà du corps. Le parapodium a deux palpes - dorsal et ventral, dans l'épiderme desquels se trouvent divers organes sensoriels qui perçoivent les irritations mécaniques et autres. Le mouvement des annélides est grandement facilité par leur division en anneaux, ce qui augmente la flexibilité du corps.
Le corps des anneaux contient des plaques compactées appelées entités frontalières, qui sous-tendent l'épiderme, séparent les muscles, sont très développés dans les cloisons entre les anneaux. Ils donnent de la force à tout le corps, servent de support au système musculo-squelettique, sont importants pour le fonctionnement des systèmes circulatoire et digestif et jouent un rôle protecteur.
Système circulatoire. Chez les annélides, en raison de la complication importante de la structure de leur corps et de l'activité fortement accrue de leurs fonctions vitales, un système de transport de substances plus avancé s'est développé - le système circulatoire. Il se compose de deux navires principaux - dorsale et ventrale(Fig. 62 et 65). Le premier passe au-dessus de l'intestin et se rapproche de ses parois, le second sous l'intestin. Dans chaque segment, les deux navires sont connectés circulaire vaisseaux. De plus, il existe des vaisseaux plus petits - ils sont particulièrement nombreux dans les parois des intestins, dans les muscles, dans la peau (à travers laquelle les gaz sont échangés), dans les cloisons séparant les segments du corps, etc. en raison de la contraction des vaisseaux eux-mêmes, principalement les vaisseaux rachidiens et annulaires antérieurs, dans les parois desquels les éléments musculaires sont bien développés.
Le sang est constitué d'une partie liquide - plasma dans lequel flottent les cellules sanguines - cellules sanguines. Le plasma contient des pigments respiratoires, c'est-à-dire des composés organiques complexes spéciaux. Ils absorbent l'oxygène dans les organes respiratoires et le libèrent dans les tissus du corps. Certaines boucles du plasma contiennent l'un des pigments respiratoires les plus avancés : l'hémoglobine ; ces anneaux ont une couleur sang rougeâtre. Le sang des annélides contient en grande partie d'autres pigments et sa couleur peut être verdâtre, jaunâtre, etc. Les cellules sanguines sont assez diverses. Parmi eux se trouvent les phagocytes qui, comme les amibes, libèrent des pseudopodes, capturent des bactéries, toutes sortes de corps étrangers, des cellules du corps mourantes et les digèrent. Comme indiqué précédemment, tous les animaux possèdent des phagocytes. Ainsi, le système circulatoire assure non seulement le transport de diverses substances, mais remplit également d'autres fonctions.
Cavité corporelle. La cavité corporelle des boucles diffère par sa structure de la cavité primaire. Cette dernière n'a pas de parois propres : à l'extérieur elle est limitée par les muscles du sac cutanéo-musculaire, à l'intérieur par la paroi intestinale (voir Fig. 53). La cavité corporelle des annélides, appelée secondaire ou coelome, est entouré d'un épithélium monocouche qui, d'une part, est adjacent au sac cutanéo-musculaire et, d'autre part, à l'intestin (voir Fig. 62). Par conséquent, la paroi intestinale devient double. L'ensemble est rempli d'un fluide aqueux, constamment en mouvement, dans lequel flottent des cellules semblables aux cellules sanguines (phagocytes, cellules à pigments respiratoires, etc.). Ainsi, la cavité corporelle secondaire, outre le rôle d’hydrosquelette, remplit des fonctions similaires à celles du sang (transfert de substances, protection contre les agents pathogènes, etc.). Cependant, il convient de souligner que le liquide coelomique se déplace plus lentement que le sang et qu'il ne peut pas entrer en contact aussi étroit avec toutes les parties du corps qu'un réseau ramifié de capillaires.
Système respiratoire. Chez les annélides, l'échange de gaz s'effectue principalement à travers la peau, mais les processus respiratoires liés à l'apparition du système circulatoire et du coelome y sont plus avancés que chez les vers précédemment considérés. De nombreuses boucles, principalement marines, ont des appendices ramifiés qui jouent le rôle de branchies (voir Fig. 61, B). La surface respiratoire augmente également en raison de la présence de diverses excroissances du corps. L'amélioration des processus respiratoires est d'une grande importance pour les annélides en raison de l'activation de leur mode de vie.
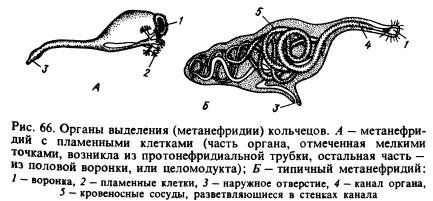
Système excréteur. Les principaux organes excréteurs sont métanéphridie(Fig. 66, B). Une métanéphridie typique consiste en un entonnoir et un long tube alambiqué, dans les parois desquels se ramifient les vaisseaux sanguins. Dans chaque segment, à l'exception de certains, on trouve deux de ces organes, à gauche et à droite de l'intestin (voir fig. 65). L'entonnoir fait face à la cavité d'un segment et le tube perce le septum, passe dans l'autre segment et s'ouvre vers l'extérieur sur la face ventrale du corps. Les produits de dissimilation sont extraits par les métanéphridies du liquide coelomique et des vaisseaux sanguins qui les entrelacent.
Chez un certain nombre d'annélides, les métanéphridies sont associées à des tubes de type protonéphridien, fermés aux extrémités tournées vers la cavité corporelle par des cellules à flamme. Il est possible que les métanéphridies soient issues de protonéphridies reliées aux entonnoirs développés sur les cloisons entre les anneaux (Fig. 66, A). On pense que ces entonnoirs, appelés coélomoductes, servait à l'origine à la sortie des produits reproducteurs de la cavité corporelle.
Sur les parois du coelome se trouvent de nombreuses cellules qui absorbent les produits de désintégration du fluide de la cavité. Il existe surtout un grand nombre de ces cellules appelées chloragogène, est présent sur les parois de la partie médiane de l’intestin. Les produits de décomposition extraits du liquide coelomique et contenus dans ces cellules ne peuvent plus avoir d'effet nocif sur l'organisme. Les cellules chargées de tels produits peuvent s’échapper par les métanéphridies ou par les pores des parois du corps.
Système digestif. Le système digestif des boucles (voir Fig. 65), en raison d'un mode de vie plus actif que dans les groupes d'animaux précédemment considérés et du progrès de l'ensemble de l'organisme, est également plus parfait. Chez les boucles : 1) la division du système digestif en différentes sections est plus prononcée, chacune remplissant sa propre fonction ; 2) la structure des parois du tube digestif est plus complexe (les glandes digestives, les muscles, etc. sont plus développés), ce qui permet de mieux traiter les aliments ; 3) l'intestin est relié au système circulatoire, grâce à quoi la digestion des nutriments et leur absorption sont plus intenses et l'apport de substances nécessaires au travail qu'il effectue est amélioré.
Le tube digestif est généralement droit et divisé en sections suivantes : cavité buccale, pharynx, œsophage, qui peut se dilater en jabot, estomac musclé (présent chez de nombreuses espèces, comme les vers de terre), intestin moyen (généralement très long), intestin postérieur. (relativement court), s'ouvrant vers l'extérieur par l'anus. Des conduits glandulaires se jettent dans le pharynx et l'œsophage, dont la sécrétion est importante dans la transformation des aliments. Chez de nombreuses boucles polychètes prédatrices, le pharynx est armé de mâchoires ; la partie antérieure du tube digestif peut se présenter sous la forme d'un tronc, ce qui permet de prendre possession de la proie et de pénétrer dans son corps. L'intestin moyen d'un certain nombre d'espèces présente une invagination profonde ( typhlosol), s'étendant sur toute la face dorsale de cet intestin (voir Fig. 62). Le typhlosol augmente la surface des intestins, ce qui accélère la digestion et l'absorption des aliments.
Reproduction. Certaines teignes se reproduisent de manière asexuée et sexuelle, tandis que d'autres ne présentent qu'une reproduction sexuée. La reproduction asexuée se fait par division. Souvent, à la suite de la division, il peut en résulter une chaîne de vers qui n'ont pas encore eu le temps de se disperser.
La structure de l'appareil reproducteur est différente. Les boucles polychètes (elles vivent dans les mers) sont dioïques et possèdent un appareil reproducteur de construction simple. Leurs gonades se développent sur les parois du coelome, les cellules germinales pénètrent dans l'eau par des fissures dans les parois du corps ou par des métanéphridies, et la fécondation des œufs a lieu dans l'eau. Les boucles vivant en eau douce et en sol humide (oligochètes), ainsi que toutes les sangsues sont hermaphrodites, leur appareil reproducteur a une structure complexe, la fécondation est interne.

Développement. L'écrasement de l'œuf fécondé, à la suite duquel les blastomères résultants sont disposés en spirale (Fig. 67), ressemble aux mêmes processus chez les vers ciliés. Les boucles polychètes se développent avec la transformation : les larves se forment à partir de leurs œufs trochophores(Fig. 68), complètement différent des vers adultes et ne se transformant en ces derniers qu'après des transformations complexes. Le trochophore est un organisme planctonique. Il est très petit, transparent et il y a généralement deux ceintures de cils le long de l'équateur de son corps : l'une, supérieure, au-dessus de la bouche, l'autre, inférieure, sous la bouche. Par conséquent, le trochophore se compose de deux parties : la partie supérieure, ou antérieure, et la partie inférieure, ou postérieure, se terminant par le lobe anal. Les trochophores de certaines espèces peuvent avoir plusieurs ceintures de cils. À l’extrémité supérieure se trouve une touffe de cils attachée à la plaque pariétale (l’organe sensoriel larvaire). Sous la plaque se trouve le centre nerveux à partir duquel s’étendent les nerfs. Le système musculaire est constitué de fibres allant dans des directions différentes. Il n'y a pas de système circulatoire. L’espace entre les parois du corps et les intestins constitue la principale cavité corporelle. Les organes excréteurs sont des protonéphridies. L'appareil digestif se compose de trois sections : antérieure, moyenne et postérieure, se terminant par l'anus. Grâce au travail des cils, la larve se déplace et la nourriture, constituée d'organismes microscopiques et de morceaux organiques, pénètre dans la bouche. Certains trochophores capturent activement les petits animaux avec leur bouche. Dans sa structure, le trochophore ressemble aux vers protocavitaires, mais à certains égards, il ressemble également aux larves de vers ciliés marins. Les parois du corps, le système nerveux, les protonéphridies, le début et la fin de l'appareil digestif, les trochophores, ont été formés à partir de l'ectoderme, la majeure partie de l'intestin - à partir de l'endoderme, les fibres musculaires - à partir de cellules appelées mésenchymateuses et provenant des deux couches .
Lorsqu’un trochophore se transforme en ver adulte, il subit un certain nombre de changements importants. Dans ces changements, le rôle le plus important est joué par les rudiments de la troisième couche germinale - mésoderme. Certains rudiments du mésoderme sont encore présents chez la larve avant le début de la métamorphose ; ils se trouvent de chaque côté entre les parois du corps et la partie postérieure de l'intestin (Fig. 68, B, 12). D'autres rudiments du mésoderme se forment plus tard à partir du bord antérieur du lobe anal, qui se transforme en zone de croissance ver (Fig. 68, B, 13). La métamorphose de la larve commence par le fait que sa partie arrière s'allonge et que les constrictions des parois du corps sont divisées en 3, 7 et rarement plus de segments. Après cela, les rudiments du mésoderme, situés entre les parois du corps et la partie postérieure de l'intestin, s'allongent également et sont divisés en autant de sections qu'il y a de segments formés à la suite de constrictions externes. Il y en a deux dans chaque anneau (Fig. 68, D, 14). Les segments formés à l'arrière du trochophore sont appelés larvaire ou larvaire, ils sont caractéristiques des derniers stades de développement du trochophore, lorsqu'il commence déjà à ressembler un peu à un ver adulte, mais comporte encore peu de segments. Au cours du développement ultérieur, des segments sont formés par la zone de croissance mentionnée ci-dessus. Ces segments sont appelés postlarvaire, ou postlarvaire(Fig. 68, D). Il y en a autant formés que le nombre de segments que possède un ver adulte d'une espèce donnée. Dans les segments postlarvaires, les rudiments mésodermiques sont d'abord divisés en sections (deux dans chaque anneau), puis le tégument externe.
Les principaux systèmes organiques d'un ver adulte sont formés comme suit (Fig. 69, A). À partir de l’ectoderme se développent l’épiderme, le système nerveux et les extrémités antérieure et postérieure du tube digestif. Les ébauches mésodermiques de chaque anneau se développent et déplacent la cavité primaire. Finalement, les rudiments droit et gauche convergent au-dessus et au-dessous de l'intestin, de sorte que le long de celui-ci, au-dessus et au-dessous, se forment des vaisseaux sanguins dorsaux et abdominaux. Par conséquent, les parois des vaisseaux sont formées à partir du mésoderme et leur cavité représente les restes de la cavité corporelle primaire. Au milieu des rudiments, les cellules s'écartent, une cavité corporelle coelomique apparaît et se développe, entourée de tous côtés par des cellules d'origine mésodermique. Cette méthode de formation du coelome est appelée téloblastique. Chaque rudiment mésodermique, en croissance, converge devant et derrière avec les rudiments voisins (Fig. 69, B) et des septa apparaissent entre eux, et les cellules mésodermiques entourant les restes de la cavité primaire entre les septa forment des vaisseaux sanguins en anneau. La couche externe des ébauches mésodermiques, adjacente à l'ectoderme, donne naissance aux muscles, la couche interne entoure le tube digestif. Par conséquent, les parois intestinales deviennent désormais doubles : la couche interne (à l'exception des extrémités antérieure et postérieure, provenant de l'ectoderme) s'est développée à partir de l'endoderme, la couche externe à partir du mésoderme. Les entonnoirs de métanéphridie sont formés à partir des cellules de la couche mésodermique et leurs tubes (représentant les restes de protonéphridies) proviennent de l'ectoderme.
Peu à peu, toutes les parties du corps d'un ver adulte se développent ; les couches de muscles se différencient, le nombre de vaisseaux sanguins augmente, l'intestin est divisé en sections, des cellules glandulaires, des fibres musculaires, des vaisseaux sanguins, etc. se développent dans ses parois. Le lobe de la tête (prostomium) d'un ver adulte est formé à partir du. partie supérieure du trochophore, l'anneau corporel des segments larvaires et postlarvaires, et le pygidium provient du lobe anal de la larve.
Origine. Diverses hypothèses ont été avancées sur l'origine des annélides. Les partisans d'une hypothèse pensent que les annélides ont évolué à partir des turbellariens. En effet, il existe des similitudes dans le développement embryonnaire des deux groupes d’animaux. Le système nerveux central des boucles (c'est-à-dire les nœuds céphaliques et la chaîne abdominale) aurait pu se former à partir du même système de turbellariens plus complexes, dans lequel les nœuds se déplaçaient vers l'extrémité antérieure du corps et deux principaux restaient du côté longitudinal. brins, et ainsi un système nerveux central de type scalène est apparu, préservé dans les annélides inférieurs. Le sac dermo-musculaire des vers plats pourrait se développer en un système d'anneaux similaire, et des métanéphridies pourraient résulter de protonéphridies. Cependant, d'un point de vue évolutif, il est impossible de supposer que les vers les plus organisés descendaient directement des vers les plus bas, chez lesquels les systèmes nerveux et musculaire étaient encore peu développés, il n'y avait pas de cavité corporelle, l'intestin n'était pas différencié. en trois sections supplémentaires et la digestion restait principalement intracellulaire, etc. d. De toute évidence, les ancêtres des vers supérieurs étaient des vers avec une structure plus complexe que les turbellariens.
Selon une autre hypothèse, les boucles seraient originaires des némertéens, c'est-à-dire des vers, descendants sans doute des turbellariens, mais présentant une structure beaucoup plus complexe que ces derniers (développement important des systèmes nerveux et musculaire, apparition d'un système circulatoire, d'un intestin traversant, etc. .). L'auteur de cette hypothèse, l'éminent zoologiste soviétique N.A. Livanov, a suggéré que dans le groupe le plus progressiste de Némertiens, des cavités métamériques sont apparues dans le sac cutané-musculaire, qui a servi de support aux muscles et s'est ensuite transformée en cavités coelomiques, comme ce qui a considérablement amélioré le mouvement des animaux. Les opposants à cette hypothèse estiment que les Némertiens, dont l'une des principales caractéristiques est un tronc absent des boucles, ne pourraient pas être les ancêtres de ces derniers. Cependant, il faut supposer que le tronc s'est développé chez les Némertiens après une longue évolution, alors qu'ils avaient des rivaux plus forts qu'auparavant dans la chasse aux animaux. Les annélides auraient pu évoluer à partir de némertes non spécialisés, dont l'organisation était déjà complexe, mais le tronc n'était pas développé. Une autre objection à l’hypothèse envisagée est plus sérieuse. De cette hypothèse, il résulte que le système circulatoire est apparu avant le coelome et que ce dernier s'est développé dès le début sous la forme de formations métamériques. Entre-temps, on connaît des vers, sans doute apparentés aux annélides, chez lesquels le métamérisme ne s'exprime pas encore, l'ensemble est continu et il n'y a pas de système circulatoire. Auparavant, on pensait que les vers mentionnés se simplifiaient en raison de leur adaptation à un mode de vie sédentaire, mais de nouvelles recherches confirment le caractère primitif originel des vers coelomiques en question.
Les auteurs de la troisième hypothèse pensent que les ancêtres des teignes étaient des vers protocavitaires, mais pas aussi spécialisés que les rotifères et les ascaris, mais plus proches des ancêtres de ce type. Cette hypothèse repose principalement sur la structure du trochophore qui, comme indiqué ci-dessus, présente d'importantes similitudes (cavité corporelle primaire, protonéphridies, via l'intestin) avec les vers protocavitaires, mais n'a toujours pas les caractéristiques des annélides. Après avoir accepté cette hypothèse, il faut supposer que le coelome est apparu à la suite du développement de l'épithélium sur les parois de la cavité corporelle primaire, et que le métamérisme corporel et le système circulatoire sont apparus plus tard. De la même hypothèse, il résulte que les Némertiens, malgré les caractéristiques progressives de leur organisation, n'étaient pas liés à l'émergence d'espèces animales plus hautement organisées. Au contraire, l'hypothèse non mertienne de l'origine des annélides rejette l'importance des vers protocavitaires pour la formation de nouveaux types d'animaux.
Il est impossible d'examiner ici de manière suffisamment détaillée les diverses objections à chacune des hypothèses mentionnées, car cela nécessite des informations plus détaillées sur la structure et le développement de tous les types de vers, mais il ne fait aucun doute que les vers coelomiques ne pourraient pas provenir directement de la vers les plus bas.
Annélides- des animaux à trois couches bilatéralement symétriques avec une cavité secondaire. Ils vivent dans les plans d’eau marins et doux, dans le sol et dans les feuilles mortes.
Le corps des annélides est divisé en un lobe céphalique, un corps segmenté et un lobe anal. Le métamérisme (répétition) des organes internes est associé à une segmentation externe. Les segments de certains annélides (polychètes) portent des membres primaires appariés - des parapodes.
Les annélides ont un sac cutanéo-musculaire constitué d'une cuticule, d'un épithélium monocouche et de deux couches de muscles (circulaires, avec des couches longitudinales en dessous). Les muscles longitudinaux sont tapissés intérieurement par un épithélium coelomique.
La cavité corporelle secondaire (coelome) est remplie de liquide coelomique, qui joue le rôle d'environnement interne du corps. Chez la plupart des espèces, le coelome est métamérique, avec des septa.
Le système circulatoire est de type fermé.
Le système digestif est divisé en trois sections fonctionnellement différentes : ectodermique antérieure, endodermique moyenne et ectodermique postérieure. Dans les parties antérieure et moyenne de l'intestin se trouvent des zones différenciées (jabot, estomac).
Les échanges gazeux se font soit à travers le tégument corporel (vers oligochètes et sangsues), soit dans les branchies (vers polychètes).
Les organes excréteurs de la plupart des espèces sont des métanéphridies, situées par paires dans chaque segment ; chez certaines espèces, des protonéphridies.
Le système nerveux comprend des ganglions suprapharyngés et sous-pharyngés appariés et une double chaîne nerveuse ventrale s'étendant à partir de ces derniers. Les ganglions suprapharyngés et sous-pharyngés sont « reliés » par des commissures en un anneau péripharyngé. La corde nerveuse ventrale est constituée de ganglions métamériquement répétés dans chaque segment.
Les annélides peuvent être dioïques ou bisexuelles. Le développement de nombreuses espèces se déroule par métamorphose. La larve des annélides marins est appelée trochophore. Certaines espèces peuvent se reproduire de manière asexuée.
Le phylum des Annélides est divisé en classes : Polychaeta, Oligochaeta, Sangsues (Hirudinea), Echiurida.
La section Le sous-règne multicellulaire à symétrie bilatérale (Bilateria) comprend également.
Type annélides rassemble environ 9 000 espèces qui ont l'organisation la plus parfaite parmi les autres vers. Leur corps est constitué d'un grand nombre de segments ; beaucoup ont des poils sur les côtés de chaque segment, qui jouent un rôle important dans le mouvement. Les organes internes sont situés dans une cavité corporelle appelée coelome. Il existe un système circulatoire. Dans la partie antérieure se trouve un groupe de cellules nerveuses qui forment les nœuds nerveux sous-pharyngés et suprapharyngés. Les annélides vivent dans les plans d’eau douce, les mers et le sol.
La plupart des représentants des annélides appartiennent aux classes : oligochètes, polychètes et sangsues.
Classe oligochètes
Représentant de la classe des oligochètes - ver de terre vit dans des terriers dans un sol humifère humide. Le ver rampe à la surface par temps humide, au crépuscule et la nuit. Chez un ver de terre, les parties antérieure et abdominale du corps se distinguent facilement. Dans la partie antérieure, il y a un épaississement de la ceinture; sur les côtés ventral et latéral du corps, des poils élastiques et courts se développent.
Le corps du ver est recouvert d'une peau constituée de tissu tégumentaire, dans laquelle les cellules adhèrent étroitement les unes aux autres. La peau contient des cellules glandulaires qui sécrètent du mucus. Sous la peau se trouvent des muscles longitudinaux circulaires et plus profonds, grâce à la contraction desquels le corps du ver peut s'allonger ou se raccourcir, se déplaçant ainsi dans le sol.
Les couches cutanées et musculaires se forment sac cutanéo-musculaire, à l'intérieur duquel se trouve une cavité corporelle où se trouvent les organes internes. Les vers de terre se nourrissent de débris végétaux en décomposition. Par la bouche et le pharynx, la nourriture pénètre dans le jabot et l'estomac musclé, où elle est broyée, pénètre dans l'intestin et y est digérée. Les substances digérées sont absorbées dans le sang et les substances non digérées sont excrétées avec la terre par l'anus.





Système circulatoire d'un ver de terre fermé et se compose de vaisseaux sanguins dorsaux et abdominaux, reliés les uns aux autres par des vaisseaux annulaires de chaque segment. Autour de l’œsophage se trouvent des vaisseaux annulaires plus grands qui font office de « cœurs » pour les branches latérales des gros vaisseaux, formant un réseau de capillaires. Le sang ne se mélange nulle part avec le liquide de la cavité corporelle, c'est pourquoi le système est appelé fermé.
Les organes excréteurs sont représentés par des tubes alambiqués à travers lesquels les liquides et les substances nocives sont éliminés du corps.
Le système nerveux est constitué de l'anneau nerveux péripharyngé et de la corde nerveuse ventrale. Le ver de terre ne possède pas d’organes sensoriels spécialisés. Il existe uniquement différents types de cellules sensibles qui perçoivent les stimuli externes (lumière, odeur, etc.).
Les vers de terre sont hermaphrodites. Cependant, ils ont une insémination croisée ; deux individus participent à ce processus. Lorsque les œufs sont pondus sur la ceinture du ver, un mucus abondant se forme dans lequel les œufs tombent, après quoi le mucus s'assombrit et durcit, formant un cocon. Ensuite, le cocon est éjecté du ver par la tête du corps. À l’intérieur du cocon, de jeunes vers se développent à partir d’œufs fécondés.
Parmi les oligochètes, il y a des nains, dont la longueur du corps ne dépasse pas quelques millimètres, mais il y a aussi des géants : Ver de terre australien 2,5 à 3 m de longueur.
Caractéristique des vers de terre capacité de régénération. Les vers de terre sont appelés formateurs de sol, car ils, en effectuant des passages dans le sol, l'ameublissent et favorisent l'aération, c'est-à-dire l'entrée de l'air dans le sol.
Polychètes de classe
Cela inclut une variété de vers marins. Parmi eux néréide. Son corps est constitué d'un grand nombre de segments. Les segments antérieurs forment la partie de la tête, sur laquelle se trouvent la bouche et les organes sensoriels : toucher - tentacules, vision - yeux. Sur les côtés du corps, chaque segment présente des lobes sur lesquels de nombreuses soies reposent en touffes. À l'aide de lames et de poils, les Néréides nagent ou se déplacent au fond de la mer. Ils se nourrissent d'algues et de petits animaux. Respirez avec toute la surface du corps. Certains polychètes ont branchies- les organes respiratoires primitifs.

Désigne les polychètes grès, vivant dans des terriers, dans le sable, ou se construisant une tortue de gypse, qui s'attache aux algues. De nombreux poissons marins se nourrissent de néréides et autres annélides.
Classe de sangsue
Le représentant le plus célèbre de cette classe est sangsue médicale, qui était déjà utilisé dans l’Antiquité pour soigner les gens. Les sangsues se caractérisent par la présence de deux ventouses : celle de devant, au fond de laquelle se trouve la bouche, et celle de derrière.
La ventouse postérieure est grande, son diamètre dépassant la moitié de la plus grande largeur du corps. Les sangsues mordent la peau avec trois mâchoires bordées de dents pointues le long des bords (jusqu'à 100 sur chaque mâchoire). Fort sangsue. En médecine, il est utilisé pour traiter les maladies des vaisseaux sanguins (formation de caillots sanguins), l'hypertension et les affections pré-AVC. Les sangsues sont appliquées sur une certaine partie d’une personne malade pour aspirer le sang. En conséquence, les caillots sanguins se dissolvent, la tension artérielle diminue et l’état de la personne s’améliore. De plus, les glandes salivaires de la sangsue médicinale produisent une substance précieuse : hirudine, - empêchant la coagulation du sang. Par conséquent, après les injections de sangsues, la plaie saigne longtemps. Dans l'estomac de la sangsue, le sang, sous l'influence de l'hirudine, se conserve pendant des mois sans être sujet à la coagulation et à la pourriture.
Le système digestif de la sangsue est conçu de telle manière qu'elle peut accumuler d'importantes réserves de sang, préservées grâce à l'hirudine. La taille d'une sangsue qui a sucé du sang augmente considérablement. Grâce à cette fonctionnalité, les sangsues peuvent mourir de faim pendant longtemps (de plusieurs mois à 1 an). Une sangsue vit jusqu'à 5 ans. Les sangsues sont hermaphrodites. Dans la nature, je réussis ! Ils n’atteignent leur maturité sexuelle qu’au cours de la troisième année de vie et pondent des cocons une fois par an en été.
Les sangsues se caractérisent par une structure droite et développée. Les sangsues comprennent la sangsue prédatrice non suceuse de sang - grand pseudokonskaya. Il se nourrit de vers (dont des sangsues), d'animaux au corps mou, de larves d'insectes aquatiques, de petits vertébrés (têtards), qu'il peut vaincre.
Les annélides sont les vers les plus organisés avec un coelome. Leurs tailles varient de quelques millimètres à 3 m. Le corps allongé est divisé en segments par des cloisons annulaires internes ; il existe parfois plusieurs centaines de segments de ce type. Chaque segment peut avoir des excroissances latérales avec des membres primitifs - des parapodes, armés de soies. La musculature est constituée de plusieurs couches de muscles longitudinaux et circulaires. La respiration s'effectue à travers la peau ; organes excréteurs - néphridies appariées, localisées segment par segment. Le système nerveux est constitué d’un « cerveau » formé de ganglions appariés et d’un cordon nerveux ventral.
Le système circulatoire fermé est constitué de vaisseaux abdominaux et dorsaux reliés dans chaque segment par de petits vaisseaux annulaires. Plusieurs des vaisseaux les plus épais de la partie antérieure du corps ont des parois musculaires épaisses et agissent comme des « cœurs ». Dans chaque segment, les vaisseaux sanguins se ramifient, formant un réseau capillaire dense.
Certaines annélides sont hermaphrodites, tandis que d’autres ont des mâles et des femelles différents. Le développement est direct ou avec métamorphose. Une reproduction asexuée (par bourgeonnement) a également lieu.
Les annélides sont divisés en 3 classes : les polychètes, les oligochètes et les sangsues.
Polychètes(Polychaeta) ont des membres primitifs (parapodes) avec de nombreuses soies sur chaque segment. Les parapodes bilobés sont souvent associés à des appendices ramifiés - des branchies, à l'aide desquelles s'effectuent les échanges gazeux. Sur la tête clairement distincte se trouvent des yeux (chez certaines espèces même capables d'accommodation), des antennes tactiles et des organes d'équilibre (statocystes). Certaines espèces sont capables de luminescence.
Pendant la saison de reproduction, les mâles libèrent du sperme dans l’eau et les femelles libèrent un grand nombre d’œufs. Chez certaines espèces, des jeux d'accouplement et une compétition pour le territoire ont été observés. La fécondation est externe ; les parents meurent alors. Le développement se produit avec métamorphose (larve nageant librement). La reproduction asexuée est rare.
Les oligochètes sont principalement des vers de sol. Parmi eux, il y a à la fois des vers de terre géants atteignant 2,5 m de long et des formes naines. Tous les segments, sauf celui oral, ont des poils disposés en touffes. Les parapodes ne sont pas prononcés, la tête est mal séparée. La fine cuticule est constamment humidifiée par le mucus sécrété ; Les échanges gazeux se font à travers la cuticule par diffusion.
Les vers oligochètes sont principalement des hermaphrodites avec fécondation croisée ; les organes génitaux sont répartis sur plusieurs segments du corps. La structure complexe de ces organes est une adaptation au mode de vie terrestre. La parthénogenèse est connue chez certaines espèces. Il n’y a pas de métamorphose ; Une douzaine de jeunes vers émergent des cocons formés lors du processus de copulation après quelques semaines.
Les sangsues (Hirudinea) ont un corps aplati, généralement de couleur brune ou verte. Il y a des ventouses aux extrémités antérieure et postérieure du corps. La longueur du corps est de 0,2 à 15 cm. Les tentacules, les parapodes et, en règle générale, les soies sont absents. Les muscles sont bien développés. La cavité corporelle secondaire est réduite. La respiration est cutanée, certains possèdent des branchies. La plupart des sangsues ont 1 à 5 paires d'yeux.
La durée de vie des sangsues est de plusieurs années. Ils sont tous hermaphrodites. Les œufs sont pondus dans des cocons ; il n'y a pas de stade larvaire. La plupart des sangsues sucent le sang de divers animaux, y compris les humains. Les sangsues percent la peau avec leur trompe ou leurs dents sur la mâchoire, et une substance spéciale - l'hirudine - empêche la coagulation du sang. Sucer le sang d'une victime peut continuer pendant des mois. Le sang dans les intestins ne se détériore pas très longtemps : les sangsues peuvent vivre même deux ans sans nourriture. Certaines sangsues sont des prédateurs et avalent leurs proies entières.
Les sangsues vivent dans les plans d’eau douce et se trouvent également dans les mers et le sol. Les sangsues servent de nourriture aux poissons. Sangsue médicale utilisé par les humains à des fins médicinales. 400 à 500 espèces.
Les annélides ont évolué à partir de vers plats primitifs du Cambrien. Les premiers annélides étaient des polychètes, qui donnèrent naissance aux oligochètes, et à travers eux, aux sangsues.
