Конечные продукты азотистого обмена азотистый обмен. Откуда берется мочевая кислота? Мочевая кислота и подагра
 Азотистый обмен - совокупность химических превращений азотсодержащих веществ в организме. А. о. включает обмен простых и сложных белков, нуклеиновых кислот, продуктов их распада (пептидов, аминокислот и нуклеотидов), содержащих азот жироподобных веществ (липидов), аминосахаров, гормонов, витаминов и др. Для нормального течения процессов жизнедеятельности организм должен быть обеспечен необходимым количеством усвояемого азота. Главнейшей составной частью и основным источником азота пищи человека являются белковые вещества
Азотистый обмен - совокупность химических превращений азотсодержащих веществ в организме. А. о. включает обмен простых и сложных белков, нуклеиновых кислот, продуктов их распада (пептидов, аминокислот и нуклеотидов), содержащих азот жироподобных веществ (липидов), аминосахаров, гормонов, витаминов и др. Для нормального течения процессов жизнедеятельности организм должен быть обеспечен необходимым количеством усвояемого азота. Главнейшей составной частью и основным источником азота пищи человека являются белковые вещества

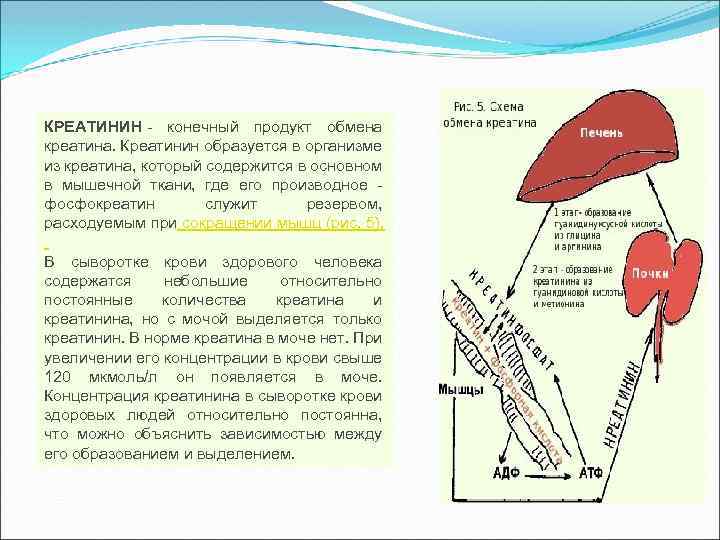 КРЕАТИНИН - конечный продукт обмена креатина. Креатинин образуется в организме из креатина, который содержится в основном в мышечной ткани, где его производное - фосфокреатин служит резервом, расходуемым при сокращении мышц (рис. 5). В сыворотке крови здорового человека содержатся небольшие относительно постоянные количества креатина и креатинина, но с мочой выделяется только креатинин. В норме креатина в моче нет. При увеличении его концентрации в крови свыше 120 мкмоль/л он появляется в моче. Концентрация креатинина в сыворотке крови здоровых людей относительно постоянна, что можно объяснить зависимостью между его образованием и выделением.
КРЕАТИНИН - конечный продукт обмена креатина. Креатинин образуется в организме из креатина, который содержится в основном в мышечной ткани, где его производное - фосфокреатин служит резервом, расходуемым при сокращении мышц (рис. 5). В сыворотке крови здорового человека содержатся небольшие относительно постоянные количества креатина и креатинина, но с мочой выделяется только креатинин. В норме креатина в моче нет. При увеличении его концентрации в крови свыше 120 мкмоль/л он появляется в моче. Концентрация креатинина в сыворотке крови здоровых людей относительно постоянна, что можно объяснить зависимостью между его образованием и выделением.
 Мочевина – конечный продукт распада белка в организме, при выведении которого через почки осуществляется удаление остатков «ненужного» азота. Образуется в печени. Выводится вместе с мочой и частично с потом (что придает поту специфический запах). Мочевина помогает организму сохранять воду и некоторые микроэлементы. Это выполняется посредством повторного всасывания почками в кровоток. Каждый раз, когда происходит процесс всасывания, мочевина «тянет» за собой обратно в кровь молекулы воды и полезных минералов. Однако избыточное ее содержание наносит вред органам и тканям.
Мочевина – конечный продукт распада белка в организме, при выведении которого через почки осуществляется удаление остатков «ненужного» азота. Образуется в печени. Выводится вместе с мочой и частично с потом (что придает поту специфический запах). Мочевина помогает организму сохранять воду и некоторые микроэлементы. Это выполняется посредством повторного всасывания почками в кровоток. Каждый раз, когда происходит процесс всасывания, мочевина «тянет» за собой обратно в кровь молекулы воды и полезных минералов. Однако избыточное ее содержание наносит вред органам и тканям.
 Мочевая кислота Конечный продукт метаболизма пуриновых оснований, входящих в состав нуклеотидов. Благодаря выведению мочевой кислоты из организма удаляется избыток азота. В плазме крови мочевая кислота содержится преимущественно в форме натриевой соли. Концентрация мочевой кислоты в крови обусловлена равновесием процессов синтеза мочевой кислоты и её выведения почками.
Мочевая кислота Конечный продукт метаболизма пуриновых оснований, входящих в состав нуклеотидов. Благодаря выведению мочевой кислоты из организма удаляется избыток азота. В плазме крови мочевая кислота содержится преимущественно в форме натриевой соли. Концентрация мочевой кислоты в крови обусловлена равновесием процессов синтеза мочевой кислоты и её выведения почками.
 ПУТИ ОБРАЗОВАНИЯ АММИАКА глутаминовая кислота -кетоглутаровая кислота + NH 3 -аминокислота -кетокислота + NH 3 цистеин пируват + NH 3 гистидин урокаиновая кислота + NH 3 глицин глиоксалевая кислота + NH 3 глюкозамин-6 -фосфат глюкоза-6 -фосфат + NH 3 глутамин глутаминовая кислота + NH 3 O ║ NH 2 -C O-P +CO 2 + NH 3 карбомоилфосфат глутамин АТФ АДФ мочевина пиримидины фолиевая кислота пурины глюкозамин
ПУТИ ОБРАЗОВАНИЯ АММИАКА глутаминовая кислота -кетоглутаровая кислота + NH 3 -аминокислота -кетокислота + NH 3 цистеин пируват + NH 3 гистидин урокаиновая кислота + NH 3 глицин глиоксалевая кислота + NH 3 глюкозамин-6 -фосфат глюкоза-6 -фосфат + NH 3 глутамин глутаминовая кислота + NH 3 O ║ NH 2 -C O-P +CO 2 + NH 3 карбомоилфосфат глутамин АТФ АДФ мочевина пиримидины фолиевая кислота пурины глюкозамин
Src="http://present5.com/presentation/229073585_437016682/image-8.jpg" alt="ПРИЧИНЫ АММИАЧНОЙ ИНТОКСИКАЦИИ ГИПЕРАММОНИЙЕМИЯ (>25 -40 мкмоль/л) Нарушение мочевинообразующей функции печени Нарушение азотовыделительной функции"> ПРИЧИНЫ АММИАЧНОЙ ИНТОКСИКАЦИИ ГИПЕРАММОНИЙЕМИЯ (>25 -40 мкмоль/л) Нарушение мочевинообразующей функции печени Нарушение азотовыделительной функции почек Острая почечная недостаточность Вирусный гепатит Хроническая почечная недостаточность Токсический гепатит Цирроз печени Портакавальные анастомозы на фоне богатой белками пищи Врожденные гипераммонийемии
 Остаточный азот и его компоненты Показатель Содержание в сыворотке крови в мг/100 мл единицы СИ Остаточный азот 20- 40 мг/100 мл 7, 06- 14, 1 ммоль/л Мочевина 20- 40 мг/100 мл 3, 3- 6, 6 ммоль/л Азот аминокислот 2, 0- 4, 3 мг/100 мл 1, 43- 3, 07 ммоль/л Мочевая кислота 2- 6, 4 мг/100 мл 0, 12- 0, 38 ммоль/л мужчины 0, 2- 0, 7 мг/100 мл 13- 53 мкмоль/л женщины 0, 4- 0, 9 мг/100 мл 27- 71 мкмоль/л мужчины 1- 2 мг/100 мл 0, 088- 0, 177 ммоль/л женщины 0, 5- 1, 6 мг/100 мл 0, 044- 0, 141 ммоль/л Аммиак 0, 03- 0, 06 мг/100 мл 21, 4- 42, 8 Креатин: Креатинин: Остальные небелковые вещества (полипептиды, нуклеотиды и др.) Ксантопротеиновая реакция 20 ед. Креатин: цельной крови 3- 4 мг % 229- 305 мкмоль/л плазмы 1- 1, 5 мг% 76, 3- 114, 5 мкмоль/л Азот мочевины крови (мочевина: 2, 14) 9- 14 мг % 3, 18- 4, 94 ммоль/л
Остаточный азот и его компоненты Показатель Содержание в сыворотке крови в мг/100 мл единицы СИ Остаточный азот 20- 40 мг/100 мл 7, 06- 14, 1 ммоль/л Мочевина 20- 40 мг/100 мл 3, 3- 6, 6 ммоль/л Азот аминокислот 2, 0- 4, 3 мг/100 мл 1, 43- 3, 07 ммоль/л Мочевая кислота 2- 6, 4 мг/100 мл 0, 12- 0, 38 ммоль/л мужчины 0, 2- 0, 7 мг/100 мл 13- 53 мкмоль/л женщины 0, 4- 0, 9 мг/100 мл 27- 71 мкмоль/л мужчины 1- 2 мг/100 мл 0, 088- 0, 177 ммоль/л женщины 0, 5- 1, 6 мг/100 мл 0, 044- 0, 141 ммоль/л Аммиак 0, 03- 0, 06 мг/100 мл 21, 4- 42, 8 Креатин: Креатинин: Остальные небелковые вещества (полипептиды, нуклеотиды и др.) Ксантопротеиновая реакция 20 ед. Креатин: цельной крови 3- 4 мг % 229- 305 мкмоль/л плазмы 1- 1, 5 мг% 76, 3- 114, 5 мкмоль/л Азот мочевины крови (мочевина: 2, 14) 9- 14 мг % 3, 18- 4, 94 ммоль/л
 гиперазотемия продукционная Печено-клеточная недостаточность немочевинных фракций остаточного азота (аммония, азота ам-т, биогенных аминов); ↓фракция мочевины в остаточном азоте Усиленный катаболизм белков (голодание, перекорм) немочевинных фракций остаточного азота (аммония, азота ам-т, биогенных аминов); фракция мочевины в остаточном азоте ретенционная Острая и хроническая почечная нед-ть ОПН: концентрация мочевины в плазме, ↓клиренс мочевины, остаточного азота и азота мочевины ХПН: остаточный азот до 200 -300 мг/100 мл, концентрация мочевины, аммония, мочевой к-ты, пептидов («средних молекул») в плазме
гиперазотемия продукционная Печено-клеточная недостаточность немочевинных фракций остаточного азота (аммония, азота ам-т, биогенных аминов); ↓фракция мочевины в остаточном азоте Усиленный катаболизм белков (голодание, перекорм) немочевинных фракций остаточного азота (аммония, азота ам-т, биогенных аминов); фракция мочевины в остаточном азоте ретенционная Острая и хроническая почечная нед-ть ОПН: концентрация мочевины в плазме, ↓клиренс мочевины, остаточного азота и азота мочевины ХПН: остаточный азот до 200 -300 мг/100 мл, концентрация мочевины, аммония, мочевой к-ты, пептидов («средних молекул») в плазме
 ИСТОЧНИКИ И СПОСОБЫ ОБЕЗВРЕЖИВАНИЯ АММИАКА В РАЗНЫХ ТКАНЯХ Биогенные амины Аминокислоты Нуклеотиды АММИАК Синтез Образование мочевины глутамина аланина глутамата аммонийных (25 г/сут) солей (0, 5 г/сут) печень мозг Мышцы, кишечник мозг почки
ИСТОЧНИКИ И СПОСОБЫ ОБЕЗВРЕЖИВАНИЯ АММИАКА В РАЗНЫХ ТКАНЯХ Биогенные амины Аминокислоты Нуклеотиды АММИАК Синтез Образование мочевины глутамина аланина глутамата аммонийных (25 г/сут) солей (0, 5 г/сут) печень мозг Мышцы, кишечник мозг почки
 РЕАКЦИИ СВЯЗЫВАНИЯ АММИАКА В КЛЕТКЕ 1. Реакция восстановительного аминирования -кетоглутарата в L-глутамат: НАДФН 2 НАДФН NH 3 + -кетоглутаровая глутаминовая + Н 2 О кислота глутаматдегидрогеназа кислота 2. Реакция образования глутамина из глутаминовой кислоты с участием фермента глутаминсинтетазы. Реакция протекает в цитозоле клеток всех тканей, но в большей степени – мозга: COOH CONH 2 │ │ CH 2 АТФ АДФ + Фн CH 2 │ │ CH 2 2 CH │ │ HC-NH 2 глутаминсинтетаза HC-NH 2 │ │ COOH COOH глутаминовая глутамин кислота
РЕАКЦИИ СВЯЗЫВАНИЯ АММИАКА В КЛЕТКЕ 1. Реакция восстановительного аминирования -кетоглутарата в L-глутамат: НАДФН 2 НАДФН NH 3 + -кетоглутаровая глутаминовая + Н 2 О кислота глутаматдегидрогеназа кислота 2. Реакция образования глутамина из глутаминовой кислоты с участием фермента глутаминсинтетазы. Реакция протекает в цитозоле клеток всех тканей, но в большей степени – мозга: COOH CONH 2 │ │ CH 2 АТФ АДФ + Фн CH 2 │ │ CH 2 2 CH │ │ HC-NH 2 глутаминсинтетаза HC-NH 2 │ │ COOH COOH глутаминовая глутамин кислота
 Пути обмена азота аминокислот и аммиака ТКАНИ КРОВЬ ПЕЧЕНЬ Мышцы, кишечник Аминок-ты -КГ Ала -КГ NH 3 Кеток-ты глутамат пируват пируват глутамат СО 2 АМФ NH 3 глюкоза карбомоил фосфат ИМФ глутамат глутамин МОЗГ и другие ткани Аминок-ты -КГ NH 3 Кеток-ты глутамат глутамин -КГ ПОЧКИ NH 3 глутамат NH 3 МОЧА орнитиновый цикл NH 3 глутамат мочевина аммонийные соли глутамат мочевина
Пути обмена азота аминокислот и аммиака ТКАНИ КРОВЬ ПЕЧЕНЬ Мышцы, кишечник Аминок-ты -КГ Ала -КГ NH 3 Кеток-ты глутамат пируват пируват глутамат СО 2 АМФ NH 3 глюкоза карбомоил фосфат ИМФ глутамат глутамин МОЗГ и другие ткани Аминок-ты -КГ NH 3 Кеток-ты глутамат глутамин -КГ ПОЧКИ NH 3 глутамат NH 3 МОЧА орнитиновый цикл NH 3 глутамат мочевина аммонийные соли глутамат мочевина
 ЦИКЛ МОЧЕВИНЫ И ЕГО СВЯЗЬ С ЦТК цитоплазма NH 3 + CO 2 + Н 2 О L-аспартат 2 АТФ карбомоил-фосфат цитруллин аргининосукцинат оксалоацетат митохондрия малат митохондрия орнитин аргинин фумарат мочевина ЦТК
ЦИКЛ МОЧЕВИНЫ И ЕГО СВЯЗЬ С ЦТК цитоплазма NH 3 + CO 2 + Н 2 О L-аспартат 2 АТФ карбомоил-фосфат цитруллин аргининосукцинат оксалоацетат митохондрия малат митохондрия орнитин аргинин фумарат мочевина ЦТК

При расщеплении белков, нуклеиновых кислот и других азотсодержащих соединений образуются токсичные вещества - аммиак, мочевина и мочевая кислота, токсический эффект которых соответственно снижается в приведенном ряду. В зависимости от того, в какой из этих трех форм преимущественно выделяется азот, животных подразделяют на три группы
:
аммониотелические
(выделяющие свободный аммиак),
уреотелические
(выделяющие мочевину) и
урикотелические
(выделяющие мочевую кислоту).
Форма выделения продуктов азотистого обмена тесно связана с условиями жизни животного и
обеспеченностью водой
. Аммиак весьма токсичен даже в малых концентрациях. Благодаря хорошей растворимости и небольшому молекулярному весу, он легко диффундирует через любую поверхность, соприкасающуюся с водой
. Аммиак
является конечным продуктом азотистого обмена
у водных беспозвоночных, костных рыб, личинок и постоянно живущих в воде земноводных.
Наземные животные ограничены в воде: чтобы избежать накопления аммиака в тканях и жидкостях тела, они должны преобразовать его в конечные продукты, нетоксичные для организма. Наземные ресничные черви, земноводные, млекопитающие выделяют мочевину.
Низкая растворимость мочевой кислоты , выпадение ее в осадок делает ее осмотически неактивной. Для ее выведения из организма вода практически не нужна. Урикотелия в основном характерна для животных, освоивших наземную, в том числе и засушливую, среду (наземные насекомые, чешуйчатые пресмыкающиеся, птицы).
Водно-солевой обмен рыб
Почки рыб выводят аммиак, соли, воду; почки наземных позвоночных - мочевину, мочевую кислоту, соли, воду. Выделительная система рыб служит для выведения из организма продуктов обмена и обеспечения его водно-солевого состава. Она включает:
Основную массу туловищной почки заполняют нефроны. Нефрон состоит из:
1) мальпигиева тельца (клубочек капиллярных сосудов, заключенный в боуменову капсулу);
2) выделительного канальца.
Артериальная кровь по почечным артериям поступает в сосудистые клубочки, где фильтруется и образуется первичная моча. В средней части выделительных канальцев происходит обратное всасывание полезных для организма веществ (сахара, витамины, аминокислоты, вода) и образуется вторичная, или окончательная, моча. У хрящевых рыб основным компонентом мочи является мочевина, у костистых - аммиак (аммиак намного токсичнее мочевины).
Выделение продуктов распада тесно связано с водно-солевым обменом рыб. У морских и пресноводных рыб эти процессы протекают различно.
When a fish eats protein, like the peel of an orange part of it goes unused and becomes waste. As Dave McShaffrey, professor of biology at Marietta College in Ohio, explains on the college website, “When proteins are converted to carbohydrates to provide energy, the amino group is removed and must be dealt with.” In saltwater fish, this nitrogen-rich waste is usually converted to either ammonia or urea, which is one of the main excretory products of saltwater fish. Ammonia is easier to produce, but urea is less toxic, requires less water and gets rid of twice as much nitrogen. Theword “ urine ” is related to “ urea .”
Морские хрящевые рыбы живут в изотоничной среде (т.е. осмотическое давление крови и тканевых жидкостей равно давлению окружающей среды). У них изотоничность внутренней и внешней среды обеспечивается за счет удержания в крови и тканевых жидкостях мочевины и солей (концентрация мочевины в крови у них достигает 2,6%). Через почки наружу у них выводятся лишь излишки мочевины, солей и воды, количество выделяемой мочи невелико (2-50 мл на 1 кг массы тела в сутки). У морских хрящевых рыб для выведения избытка солей сформировалась особая ректальная железа, открывающаяся в прямую кишку.
Все пресноводные рыбы живут в гипотонической среде (т.е. осмотическое давление крови и тканевых жидкостей выше, чем в окружающей среде), поэтому вода постоянно проникает в организм через кожу, жабры, с пищей. Чтобы избежать обводнения пресноводные рыбы имеют хорошо развитый фильтрационный аппарат почек, что позволяет им выделять большое количество мочи (50-300 л на 1 кг массы тела в сутки). Потеря солей с мочой компенсируется активной их реабсорбцией в почечных канальцах и поглощением солей жабрами из воды, часть солей поступает с пищей.
Морские костистые рыбы живут в гипертонической среде (т.е. осмотическое давление крови и тканевых жидкостей ниже, чем в окружающей среде), поэтому вода выходит из организма через кожу, жабры, с мочой и фекалиями. Во избежание иссушения они пьют соленую воду (от 40 до 200 мл на 1 кг массы в сутки), которая из кишечника всасывается в кровь. У морских костистых рыб уменьшается число клубочков в почках, а у некоторых рыб исчезают совсем (морская игла, морской черт). Таким образом, почки выводят небольшое количество мочи (0,5-20 мл на 1 кг массы тела в сутки).
Проходные рыбы при переходе из одной среды в другую могут изменять способ осморегуляции: в морской среде она осуществляется как у морских рыб, а в пресной - как у пресноводных. Такие адаптации водно-солевого обмена позволили костистым рыбам широко освоить пресные и соленые водоемы.
Адаптации наземных животных к выделению веществ
According to New World Encyclopedia, reptiles use two small kidneys as tools for excretion. The kidneys serve to filter the nitrogen from the animal"s bloodstream, then turn it into waste. The nitrogen then exits the body in dry form as uric acid crystals along with the feces. According to Stanford University, the kidneys in a bird also function as a means to remove nitrogen from the blood. The white substance found in bird droppings is actually uric acid, which is not water soluble. In both reptiles and birds, eliminating the nitrogen requires that the body exerts a great deal of energy. Both species are able to efficiently remove the nitrogen while losing very little water in the waste product.
Адаптации пустынных животных к выделению веществ
Обитатели полупустынных биотопов большую часть воды получают, поедая сочные части суккулентных растений. Их кожно-легочные потери воды минимальны. Так, при температуре 20°С они достигают у относительно влаголюбивого вида - гребенщиковой песчанки 170 см
3
, а у сухолюбивой большой песчанки - только 50 см
3
на 1 кг массы за 1 ч. Настоящие пустынные млекопитающие способны поедать почти сухие корма и практически не пить в течение всей жизни, удовлетворяя свои потребности лишь за счет образующейся в организме метаболической воды. Верблюды в кормные и влажные сезоны запасают жир, расходуемый в малокормное и сухое время - при этом образуется некоторое количество воды; наконец, во время отдыха и сна они снижают температуру тела, что также сокращает расход воды.
Desert Animals
Coping with water loss is a particular problem for animals that live in dry conditions. Some, like the camel, have developed great tolerance for dehydration. For example, under some conditions, camels can withstand the loss of one third of their body mass as water. They can also survive wide daily changes in temperature. This means they do not have to use large quantities of water in sweat to cool the body by evaporation.Smaller animals are more able than large ones to avoid extremes of temperature or dry conditions by resting in sheltered more humid situations during the day and being active only at night.The kangaroo rat is able to survive without access to any drinking water at all because it does not sweat and produces extremely concentrated urine. Water from its food and from chemical processes is sufficient to supply all its requirements.


Какой из нефронов принадлежит верблюду, а какой – рептилии? Почему вы сделали такой выбор?
Fresh Water Fish
Although the skin of fish is more or less waterproof, the gills are very porous. The body fluids of fish that live in fresh water have a higher concentration of dissolved substances than the water in which they swim. In other words the body fluids of fresh water fish are hypertonic to the water (see chapter 3). Water therefore flows into the body by osmosis . To stop the body fluids being constantly diluted fresh water fish produce large quantities of dilute urine.
Marine Fish
Marine fish like the sharks and dogfish have body fluids that have the same concentration of dissolved substances as the water (isotonic ) have little problem with water balance. However, marine bony fish like red cod, snapper and sole, have body fluids with a lower concentration of dissolved substances than seawater (they are hypotonic to seawater). This means that water tends to flow out of their bodies by osmosis. To make up this fluid loss they drink seawater and get rid of the excess salt by excreting it from the gills.
Marine Birds
Marine birds that eat marine fish take in large quantities of salt and some only have access to seawater for drinking. Bird’s kidneys are unable to produce very concentrated urine, so they have developed a salt gland. This excretes a concentrated salt solution into the nose to get rid of the excess salt.
2. Using the words/phrases in the list below fill in the blanks in the following statements.
| cortex | amino acids | renal | glucose | water reabsorption | large proteins |
| bowman’s capsule | diabetes mellitus | secreted | antidiuretic hormone (ADH) | blood cells |
| glomerulus | concentration of the urine | medulla | nephron |
a) Blood enters the kidney via the ......................... artery.
b) When cut across the kidney is seen to consist of two regions, the outer.............. and the inner..............
c) Another word for the kidney tubule is the...............................
d) Filtration of the blood occurs in the..............................
e) The filtered fluid (filtrate) enters the.............................
f) The filtrate entering the e) above is similar to blood but does not contain.................. or....................
g) As the fluid passes along the first coiled part of the kidney tubule.................. and.................... are removed.
h) The main function of the loop of Henle is.............................................................
i) Hydrogen and potassium ions are.............................. into the second coiled part of the tubule.
j) The main function of the collecting tube is..................................................
k) The hormone...................................... is responsible for controlling water reabsorption in the collecting tube.
l) When the pancreas secretes inadequate amounts of the hormone insulin the condition known as............................... results. This is most easily diagnosed by testing for................................ in the urine.
Форма выведения белкового азота - в виде аммиака, мочевины или же мочевой кислоты - тесно связана с условиями жизни цветного и наличием воды (табл. 10.4). Аммиак весьма токсичен даже в очень малых концентрациях, поэтому он должен быстро
удаляться либо путем выведения во внешнюю среду, либо путем превращения в менее токсичные вещества (мочевину или мочевую кислоту).
У большинства водных беспозвоночных конечным продуктом белкового обмена является аммиак. Благодаря его легкой растворимости и небольшому молекулярному весу он диффундирует чрезвычайно быстро. Значительная его часть может быть выделена через любую поверхность, соприкасающуюся с водой, - не обязательно через почку. У костистых рыб большая часть азота выводится в форме аммиака через жабры. У карпа и золотой рыбки жабры выделяют в 6-10 раз больше азота, чем почки, и только 10% его составляет мочевина; остальные 90% выводятся в виде аммиака (Smith, 1929).
МОЧЕВИНА
Мочевина легко растворима в воде и обладает довольно малой токсичностью. Синтез мочевины у высших животных был изучен знаменитым биохимиком Гансом Кребсом - тем самым ученым, по имени которого был назван цикл окислительного энергетического обмена (цикл трикарбоновых кислот, или цикл Кребса),
При синтезе мочевины аммиак и двуокись углерода, конденсируясь с фосфатом, образуют карбамоилфосфат, который затем используется для синтеза цитруллина из орнитина, как показано на рис. 10.13. После этого добавляется еще одна молекула аммиака из аспарагиновой кислоты, и это ведет к образованию аминокислоты аргинина. В присутствии фермента аргиназы аргинин распадается на мочевину и орнитин. Из орнитина синтезируется новая молекула цитруллина, и весь цикл повторяется; поэтому весь этот путь превращений называют орнитиновым циклом синтеза мочевины. Наличие аргиназы у животного говорит о его способности вырабатывать мочевину и часто указывает на то, что мочевина является у него главным азотистым экскретом. Но это необязательно так, поскольку возможно наличие аргиназы и при: отсутствии всего цикла.
МОЧЕВИНА У ПОЗВОНОЧНЫХ
Позвоночные животные, выделяющие главным образом мочевину и обладающие для ее синтеза ферментами орнитинового цикла, представлены на рис. 10.14. Некоторое количество мочевины выделяют костистые рыбы, а у пластиножаберных, амфибий и млекопитающих это главный азотистый экскрет. У пластиножаберных (акул и скатов), а также у крабоядной лягушки и целаканта Latimeria мочевина задерживается в организме, играет
важную роль в саморегуляции и поэтому является ценным продуктом обмена. У пластиножаберных мочевина фильтруется в почечном клубочке, но ввиду ее значения для осморегуляции она не должна теряться с мочой; поэтому она возвращается в результате активной реабсорбции в канальцах. У амфибий дело обстоит иначе.

Мочевина фильтруется, и, кроме того, значительное количество ее добавляется к моче путем активной секреции в канальцах. Таким образом, и у пластиножаберных, и у амфибий имеет место активный канальцевый транспорт мочевины, но он идет у этих групп в разных направлениях. Очевидно, насосные механизмы здесь метаболически не идентичны, поскольку опыты с рядом близких друг к другу производных мочевины дают в обеих группаx животных различные результаты (табл. 10.5). Это превосходный пример того, как одна и та же физиологическая функция возникает в двух группах независимо, причем для достижения одной и той же цели (в данном случае для активного транспорта мочевины) не обязательно используются одинаковые механизмы.
У крабоядной лягушки, которая тоже сохраняет мочевину для осморегуляции, активной реабсорбции этого вещества в канальцах
не обнаружено (Schmidt-Nielsen, Lee, 1962). Моча образуется у нее медленно, и почечные канальцы весьма проницаемы для мочевины. Поэтому мочевина диффундирует из канальцевой жидкости

Рис. 10.14. Выделение азота на разных этапах филогенеза позвоночных. Линиям" окружены группы животных, которые выделяют соответственно аммиак, мочевину и мочевую кислоту в качестве основного экскрета. (В. Schmidt-Nielsen, 3972.)
обратно в кровь и оказывается в моче приблизительно в той же концентрации, что и в крови. Таким образом, лишь небольшие количества ее теряются с мочой.
Если у обычных лягушек происходит активная канальцевая секреция мочевины, то почему крабоядная лягушка не использует
Таблица 10.5
Мочевина активно транспортируется почечным канальцем акулы (активная реабсорбция) и лягушки (активная секреция). Но с тремя другими близкими веществами результаты получаются у этих двух видов животных совершенно различными. Это говорит о том, что клеточный механизм транспорта в их почках различен. (В. Schmidt-Nielsen, Rabinovitz, 1964)

такой насос, попросту поменяв его направление на обратное? На этот вопрос ответить нелегко, но, по-видимому, направление активного транспорта - консервативная физиологическая функция, изменить которую непросто. Как мы уже видели, и в коже лягушки, и в почке млекопитающего сохраняется направление активного транспорта хлористого натрия извне внутрь организма. Но в почке млекопитающего направленный внутрь обратный транспорт NaCl из канальцевой жидкости в организм используется в умножающей противоточной системе таким образом, что конечным результатом тем не менее оказывается концентрированная моча.
Обычное представление о выделении мочевины почкой млекопитающего состоит в том, что мочевина фильтруется в клубочке, а затем пассивно проходит по канальцам, хотя некоторая ее доля благодаря высокой способности к диффузии пассивно диффундирует обратно в кровь. Имеются, однако; убедительные доказательства того, что мочевина служит важным элементом умножающей противоточной системы и что способ выделения мочевины является существенным элементом функции почки у млекопитающих.
МОЧЕВИНА И МЕТАМОРФОЗ У АМФИБИЙ
Головастики лягушек и жаб выделяют главным образом аммиак; взрослые животные выделяют мочевину. У лягушки (Rana temporaria), жабы (Bufo bufo), тритона (Triturus uulgaris) и других амфибий при метаморфозе происходит четкий переход от выделения аммиака к экскреции мочевины. Однако южноафриканская шпорцевая лягушка (Xenopus), которая и во взрослом состоянии остается в воде, продолжает выделять аммиак и на этой: :тадии (табл. 10.6).
Переход к выделению мочевины во время метаморфоза у полуназемных амфибий связан; с заметным повышением активности всех ферментов орнитинового цикла в печени (Brown et al., 1959).
Таблица 10.6
Выделение аммиака у наземной жабы Bufo bufo и у полностью водной бесхвостой амфибии Xenopus laevis. Цифры указывают выделение свободного аммиака в процентах от общего количества выделяемых аммиака и мочевины на разных стадиях развития. (Munro, 1953 )

Интересно, что особи водной амфибии Xenopus, извлеченные:на несколько недель из воды, накапливают в крови и тканях мочевину. Накопление мочевины можно вызвать, поместив животных в 0,9%-ный раствор NaCl. Когда взрослых особей содержали вне воды, но в сыром мху (чтобы избежать обезвоживания), концентрация мочевины в крови увеличивалась в 10-.20 раз и достигала почти 100 ммоль/л. После возвращения животных в воду избыточная мочевина выделялась (Balinsky et al, 1961).
У группы особей Xenopus, которые в естественных условиях переживали летнюю засуху в иле около высохшего пруда, концентрация мочевины также была повышена в 15-20 раз. Среди ферментов, участвующих в синтезе мочевины, количество карбамоилфосфатсинтетазы, ответственной за первый этап синтеза (см. рис. 10.13), возросло приблизительно в шесть раз, но активность остальных ферментов цикла не изменилась. Возможно, что синтез карбамоилфосфата является этапом, лимитирующим скорость синтеза мочевины, и увеличение количества этого фермента, вероятно, удерживает аммиак плазмы на низком уровне, когда животные находятся вне воды (Balinsky et al., 1967).
МОЧЕВИНА У ДВОЯКОДЫШАЩИХ РЫБ
У африканской двоякодышащей рыбы Protopterus происходят совершенно такие же изменения, как у амфибий. В обычных условиях, когда такая рыба живет в воде, она выделяет много аммиака
(и некоторое количество мочевины), но когда в период засухи она находится в коконе в засохшем иле, то все азотистые отходы превращаются у нее в мочевину, которая накапливается в крови, где ее концентрация к концу трехлетнего пребывания рыбы в коконе может достигать 3% (500 ммоль/л) (Smith, 1959).
В печени африканской двоякодышащей рыбы обнаружены все пять ферментов орнитинового цикла (Janssens, Cohen, 1966). Уровни двух ферментов, лимитирующих скорость синтеза мочевины, сходны у этой рыбы и у головастика лягушки Rana catesbeiaпа и значительно ниже уровней, найденных у взрослых лягушек. Это согласуется с фактом преимущественного выделения аммиака двоякодышащей рыбой, когда она находится в воде. Однако было вычислено, что того количества ферментов орнитинового цикла, которое содержится в печени двоякодышащей рыбы, не находящейся в спячке, достаточно, чтобы обеспечить накопление мочевины, фактически наблюдаемое во время спячки (Forster, Goldstein, 1966).
У австралийской двоякодышащей рыбы Neoceratodus концентрации ферментов орнитинового цикла невелики, что согласуется с образом жизни этой рыбы: она пользуется легким только как: добавочным органом дыхания и может лишь недолго выживать на воздухе (о дыхании двоякодышащих рыб см. гл. 2). Синтез мочевины в срезах печени австралийской двоякодышащей рыбы идет в сто раз медленнее, чем у африканской. Это опять-таки согласуется с чисто водным образом жизни первой из них (Goldstein et al., 1967).
МОЧЕВАЯ КИСЛОТА
Выделение мочевой кислоты преобладает у насекомых, наземных улиток, большинства рептилий и у птиц. Все это - типично наземные животные, и образование у них мочевой кислоты можно рассматривать как эффективное приспособление, сберегающее воду при жизни на суше. Поскольку мочевая кислота и ее соли очень плохо растворимы в воде (растворимость ее составляет около 6 мг на 1 л воды), реабсорбция воды из мочи ведет к выпадению мочевой кислоты и ее солей в осадок.
МОЧЕВАЯ КИСЛОТА У ПТИЦ И НАСЕКОМЫХ
Полутвердая белая часть птичьего помета представляет собой мочу и состоит главным образом из мочевой кислоты; для выведения азотистых экскретов птицы расходуют очень мало воды, некоторых насекомых уменьшение потерь воды с мочой зашло так далеко, что они вообще не выделяют мочевую кислоту, а откладывают ее в разных частях организма, главным образом в
жировом теле. Поэтому для удаления конечных азотистых продуктов таким формам вода совсем не нужна (Kilby, 1963).
Было высказано предположение, что использование мочевой кислоты в качестве главного экскрета дает птицам еще одно преимущество. Поскольку для образования мочи им нужно мало воды, выделение мочевой кислоты, согласно этому предположению, уменьшает вес тела у летающих птиц. Но это соображение не убедительно, так как птицы, имеющие доступ к воде (как пресноводные, так и морские), часто выделяют большие количества жидкой мочи.
КЛЕЙДОИЧЕСКОЕ ЯЙЦО
Джозеф Нидхем предположил, что разница между теми позвоночными, которые вырабатывают мочевину (млекопитающие и амфибии), и теми, которые продуцируют мочевую кислоту (рептилии и птицы), связана прежде всего со способом размножения. Яйцо амфибии развивается в воде, а эмбрион млекопитающего - в жидкой среде в матке, где отходы метаболизма попадают в кровь матери. С другой стороны, эмбриональное развитие рептилий и птиц происходит в замкнутом, так называемом клейдоическом яйце, которое обменивается с внешней средой только газами, а все экскреты остаются внутри скорлупы. В клейдоическом яйце запас воды очень невелик, а аммиак, разумеется, слишком токсичен, чтобы эмбрион мог выносить его присутствие в больших количествах. Если бы вырабатывалась мочевина, она оставалась бы в яйце и накапливалась в растворенном состоянии. Между тем мочевая кислота может выпадать в осадок и тем самым по существу элиминироваться; это и происходит, когда она откладывается в виде кристаллов в аллантоисе, который, таким образом, служит эмбриональным мочевым пузырем.
МОЧЕВАЯ КИСЛОТА У РЕПТИЛИЙ
Ящерицы и змеи выделяют главным образом мочевую кислоту; многие черепахи выделяют смесь мочевой кислоты и мочевины, а крокодилы - главным образом аммиак (Cragg et al., 1961). Это соответствует общему представлению, что способ экскреции азота тесно связан с количеством доступной воды в окружающей среде.
Крокодилы и аллигаторы выделяют аммиак в моче, где главным катионом является NH4+, а главным анионом - НСО 3 - (Со-ulson et al., 1950; Goulson, Hernandez, 1955). Возможно, что присутствие в моче указанных ионов помогает этим пресноводным
животным лучше удерживать ионы Na + и С1 - , потеря которых с калом, кстати, тоже очень невелика.
Вряд ли можно сомневаться в тесной связи между средой обитания черепах и выделением ими азота. В табл. 10.7 приведен состав проб мочи от восьми видов черепах, полученных из Лондонского зоопарка. У видов с наиболее выраженным водным образом жизни выделяются значительные количества аммиака и мочевины и только следы мочевой кислоты; у наиболее сухопутных форм больше половины азота выводится в виде мочевой кислоты.
Таблица 10.7
Доля азота в моче различных черепах (в процентах от всего выделяемого азота). Формы, в наибольшей степени связанные с водой, почти не выделяют мочевой кислоты, но это вещество доминирует у наземных видов из засушливых областей. Moyle, 1949 )

Сведения о том, выделяют ли черепахи главным образом мочевину или же мочевую кислоту, противоречивы. Дело в том, что различаются не только виды, но и внутри одного вида одни особи могут выделять преимущественно мочевую кислоту, другие - преимущественно мочевину, третьи - смесь обоих веществ Khalil, Haggag, 1955). Даже одна и та же особь может с течением времени перейти от одного соединения к другому. Некоторое
количество выпавшей в осадок мочевой кислоты задерживается в клоаке, а жидкая часть мочи выводится наружу; это делает ненадежным определение образующейся мочевой кислоты путем анализа одной или нескольких проб мочи: при неполном опорожнении клоаки могут получиться совсем низкие цифры, а при такой ее эвакуации, когда выходит осадок, накопившийся за некоторое время, мочевой кислоты окажется слишком много.
У черепахи Testudo mauritanica переход от мочевины к мочевой кислоте и обратно находится, по-видимому, в прямой зависимости от температуры и содержания воды в организме. Выделение мочевой кислоты возрастает при неблагоприятном водном балансе, но механизм, управляющий этим сдвигом биохимической активности, неясен.
В главе 9 мы уже упоминали, что африканская лягушка Chiromantis xerampelina теряет воду через кожу очень медленно, примерно с той же скоростью, что и рептилии. Она сходна с рептилиями и тем, что выделяет в основном мочевую кислоту, а не мочевину, как это обычно свойственно взрослым амфибиям. Эта сенсационный факт, так как он противоречит общепринятому представлению о выделении азота у амфибий. Точность этого сообщения не вызывает сомнений, так как мочевую кислоту определяли в моче Chiromantis специфичным для этого вещества ферментативным методом, и было установлено, что она составляет до 60-75% сухого веса мочи (Loveridge, 1970).
Южноафриканская лягушка Phyllotnedusa sauvagii в этом отношении тоже сходна с рептилиями. Потеря воды через кожу составляет у нее величину того же порядка, что и у рептилий с их сухими покровами, а моча содержит много полутвердого осадка урата (Shoemaker et al., 1972). В форме урата у Phyllomedusa выделяется 80% общего азота, и повышенное потребление воды не изменяет интенсивности образования урата. Этот вид продолжает выделять главным образом мочевую кислоту даже при избытке воды. Когда лягушка нуждается в сохранении воды, экскреция мочевой кислоты (вместо мочевины) приобретает очень большое значение. Вычислено, что если бы экскреторным продуктом у этой лягушки была мочевина, то для образования мочи ей потребовалось бы около 60 мл воды в день на 1 кг веса тела. А между тем, благодаря тому что P. sauvagii выделяет мочевую кислоту, она теряет с мочой всего лишь 3,8 мл воды в день на 1 кг веса тела (Shoemaker, McClanahan, 1975).
АММИАК И ПОЧЕЧНАЯ ФУНКЦИЯ
Из всего сказанного выше может создаться впечатление, что аммиак выделяют главным образом водные животные, но это не совсем верно. Аммиак в норме содержится и в моче наземных животных, где он служит для регуляции рН мочи. Если моча становится кислой из-за выделения кислых продуктов обмена, для нейтрализации добавляется аммиак.
Избыток кислоты обычно образуется при белковом обмене, так как конечным продуктом окисления серусодержащей аминокислоты цистеина является серная кислота. Чем кислее моча, тем больше добавляется аммиака. Аммиак, используемый для нейтрализации кислой мочи, образуется в почках из аминокислоты глутамина. Почки содержат глутаминазу, и она имеется здесь специально для выработки аммиака. Поэтому аммиак в моче млекопитающего прямо не связан с тем аммиаком, который образуется в печени при дезаминировании аминокислот, и в этом смысле его не следует рассматривать как нормальный конечный продукт белкового обмена.
НУКЛЕИНОВЫЕ КИСЛОТЫ И ВЫДЕЛЕНИЕ АЗОТА
Нуклеиновые кислоты содержат две группы азотистых соединений: пурины (аденин и гуанин) и пиримидины (цитозин и тимин). У некоторых животных пурины выделяются в виде мочевой кислоты (которая и сама является пурином); у других животных пуриновая структура расщепляется до ряда промежуточных соединений или до аммиака, причем любое из этих веществ может выводиться из организма.
Метаболическое расщепление пуринов и выделение его конечных продуктов изучены не так тщательно, как обмен белкового 13ота. Важнейшие данные приведены в табл. 10.8. У птиц, наземных рептилий и насекомых пурины расщепляются до мочевой кислоты и последняя выводится из организма. Это те животные, у которых из аминного азота синтезируется мочевая кислота; очевидно, что для животного было бы бессмысленно синтезировать мочевую кислоту и в то же время обладать механизмами ее разложения. Поэтому нельзя ожидать дальнейшего распада пуринов у тех животных, у которых мочевая кислота - конечный продукт белкового обмена.
Среди млекопитающих человек, высшие обезьяны и далматский дог составляют особую группу: они выделяют мочевую кислоту, тогда как остальные млекопитающие выделяют аллантоин. Аллантоин образуется из мочевой кислоты путем одного превращения в присутствии фермента уриказы. У человека и высших обезьян нет этого фермента. Из-за своей малой растворимости мочевая кислота иногда откладывается в организме человека, вызывая припухлость суставов и очень мучительное заболевание -
Таблица 10.8
Азотистые конечные продукты пуринового обмена у разных животных. (Keilin, 1959)

подагру. Если бы у человека сохранился фермент уриказа, подагры не существовало бы.
Хотя далматский дог выделяет гораздо больше мочевой кислоты, чем другие собаки, это не следствие какого-то дефекта метаболизма. Печень всякой собаки содержит уриказу и вырабатывает некоторое количество аллантоина. Но у далматского дога имеется почечный дефект, препятствующий канальцевой реабсорбции мочевой кислоты (которая происходит у других млекопитающих, в том числе у человека); поэтому у дога мочевая кислота теряется с мочой быстрее, чем перерабатывается печенью в аллантоин (Yu et al., I960). Немало данных говорит о том, что мочевая кислота у далматского дога не только фильтруется в клубочке, но и экскретируется путем активного транспорта в канальцах (Keilin, 1959).
Пурины аденин и гуанин сходны по своей структуре с мочевой кислотой: они содержат одно шестичленное кольцо и одно пятичленное. Но пиримидины (цитозин и тимин) представляют собой одиночные шестичленные кольца, содержащие два атома азота. У высших позвоночных пиримидины расщепляются путем разрыва этого кольца с образованием одной молекулы аммиака и одной молекулы β-аминокислоты. Последняя метаболизируется затем по обычной схеме дезаминирования.
Самая поразительная черта обмена нуклеиновых кислот состоит в том, что "высшие" животные, перечисленные в начале табл. 10.8, полностью лишены ферментов, нужных для расщепления
пуринов. Среди "низших" животных мы находим все большее усложнение биохимических и ферментных систем, осуществляющих дальнейшее расщепление пуринов, так что самые "низшие" формы обладают наиболее полным ферментным аппаратом.
ДРУГИЕ АЗОТИСТЫЕ СОЕДИНЕНИЯ
У пауков главным экскретом является гуанин. По-видимому, он синтезируется из аминного азота, хотя весь путь его образования неизвестен. Некоторые пауки, в том числе птицеядные тарантулы, после приема пищи выделяют более 90% всего азота в форме гуанина (Peschen, 1939). У обыкновенного садового паука Epeira diadema идентификация гуанина была подтверждена весьма специфическим ферментативным методом (Vajropala, 1935).
Гуанин довольно часто встречается и у разнообразных других животных. Например, серебристый блеск рыбьей чешуи обусловлен отложением кристаллов гуанина. Садовая улитка Helix выделяет гуанин, но лишь в пределах около 20% общего количества экскретируемых пуринов, а остальные 80% составляет мочевая кислота. Возможно, что эта фракция - продукт обмена нуклеиновых кислот, а мочевая кислота образуется в результате белкового обмена.
Аминокислоты не занимают важного места среди продуктов азотистого обмена, но в небольших количествах они содержатся в моче многих животных. Казалось бы, животному выгоднее дезаминировать аминокислоту, выделять аммиак обычным путем и использовать образующуюся органическую кислоту в энергетическом обмене. Но поскольку аминокислоты играют лишь незначительную роль в выделении азота, этот вопрос не будет здесь обсуждаться.
ТЕОРИЯ РЕКАПИТУЛЯЦИИ
Обычно считали, что выделение азота у развивающегося куриного эмбриона изменяется во времени и проходит через ряд пиков: вначале основным продуктом является аммиак, затем мочевина и, наконец, мочевая кислота. Предполагалось, что такое развитие рекапитулирует этапы эволюции, которая у птиц заканчивается выделением мочевой кислоты. Как сообщалось, образование аммиака у куриного зародыша достигает максимума через 4 дня, мочевины - через 9 дней и мочевой кислоты - через 11 дней после начала инкубации (Baldwin, 1949).
Более новые работы говорят о том, что выделение азота у куриного эмбриона резко отличается от этой ранее описанной картины (Clark, Fischer, 1957). Все три главных экскреторных продукта- аммиак, мочевина и мочевая кислота - образуются и присутствуют с самого начала эмбрионального развития. К концу
периода инкубации мочевой кислоты становится намного больше, чем остальных двух продуктов. Однако количество мочевины и аммиака продолжает расти на протяжении всей инкубации, и ко времени вылупления оба вещества содержатся примерно в одинаковых количествах. К концу инкубации количество выделяемого азота достигает 40 мг, из которых 23% делятся поровну между мочевиной и аммиаком, а остальное представлено мочевой кислотой (рис. 10.15).

В чем причина расхождения в полученных результатах? Прежние данные могли быть менее точными из-за более примитивных методов анализа, но этим вряд ли можно объяснить наблюдавшиеся отдельные пики. Главная причина состоит просто в том, что результаты выражались в количествах каждого экскреторного продукта на единицу веса зародыша. А поскольку зародыш непрерывно и чем дальше, тем быстрее увеличивается в размерах,
то при делении количества каждого вещества на вес эмбриона создается искусственный пик.
На самом деле все три экскреторных продукта присутствуют с самого начала и на протяжении эмбрионального развития их становится постепенно все больше, но после 10-го дня инкубации количество аммиака возрастает незначительно. Мочевина, вырабатываемая зародышем, синтезируется не из азота аминокислот в цикле орнитина, а в результате воздействия аргиназы на аргинин (Eakin, Fisher, 1958). Таким образом, ни образование аммиака, ни синтез мочевины в курином эмбрионе не подтверждают представление о том, что онтогенез биохимических механизмов повторяет эволюционную историю выделения азота.
Мы рассмотрели разнообразные органы выделения и описали их общие особенности. Эти органы удаляют отходы метаболизма, помогают поддерживать нужные концентрации солей и других растворенных веществ и регулируют содержание воды в организме, тщательно сохраняя воду, если ее в организме мало, и выводя ее избыточные количества.
Убедительные данные показывают, однако, что это не всегда верно, Некоторые исследователи (например, Costa et al., 1968, 1974) сообщают об образовании газообразного азота у млекопитающих, получающих большие количества белка. Эти сведения должны изменить некоторые из наших представлений о белковом обмене и конечных азотистых продуктах.
По-гречески kleisto - замкнутый, от kleis - ключ.
Два описанных здесь вида лягушек обитают в сухих, полупустынных местностях. - Прим. ред.
Азотистый обмен - совокупность химических превращений, реакций синтеза и распада азотистых соединений в организме; составная часть обмена веществ и энергии. Понятие «азотистый обмен» включает в себя белковый обмен (совокупность химических превращений в организме белков и продуктов их метаболизма), а также обмен пептидов, аминокислот , нуклеиновых кислот , нуклеотидов, азотистых оснований, аминосахаров (см. Углеводы), азотсодержащих липидов , витаминов , гормонов и других соединений, содержащих азот.
Организм животных и человека усвояемый азот получает с пищей, в которой основным источником азотистых соединений являются белки животного и растительного происхождения. Главным фактором поддержания азотистого равновесия - состояния А. о., при котором количество вводимого и выводимого азота одинаково, - служит адекватное поступление белка с пищей. В СССР суточная норма белка в питании взрослого человека принята равной 100 г , или 16 г азота белка, при расходе энергии 2500 ккал . Азотистый баланс (разность между количеством азота, который попадает в организм с пищей, и количеством азота, выводимого из организма с мочой, калом, потом) является показателем интенсивности А. о. в организме. Голодание или недостаточное по азоту питание приводят к отрицательному азотистому балансу, или азотистому дефициту, при котором количество азота, выводимого из организма, превышает количество азота, поступающего в организм с пищей. Положительный азотистый баланс, при котором вводимое с пищей количество азота превышает количество азота, выводимое из организма, наблюдается в период роста организма, при процессах регенерации тканей и т.д. Состояние А. о. в значительной степени зависит от качества пищевого белка, которое, в свою очередь, определяется его аминокислотным составом и прежде всего наличием незаменимых аминокислот.
Принято считать, что у человека и позвоночных животных А. о. начинается с переваривания азотистых соединений пищи в желудочно-кишечном тракте. В желудке происходит расщепление белков при участии пищеварительных протеолитических ферментов трипсина и гастриксина (см. Протеолиз ) с образованием ептидов, олигопептидов и отдельных аминокислот. Из желудка пищевая масса поступает в двенадцатиперстную кишку и нижележащие отделы тонкой кишки, где пептиды подвергаются дальнейшему расщеплению, катализируемому ферментами сока поджелудочной железы трипсином, химотрипсином и карбоксипептидазой и ферментами кишечного сока аминопептидазами и дипептидазами (см. Ферменты ). Наряду с пептидами. в тонкой кишке расщепляются сложные белки (например, нуклеопротеины) и нуклеиновые кислоты. Существенный вклад в расщепление азотсодержащих биополимеров вносит и микрофлора кишечника. Олигопептиды, аминокислоты, нуклеотиды, нуклеозиды и др. всасываются в тонкой кишке, поступают в кровь и с ней разносятся по всему организму. Белки тканей организма в процессе постоянного обновления также подвергаются протеолизу под действием тканевых протсаз (пептидаз и катепсинов), а продукты распада тканевых белков попадают в кровь. Аминокислоты могут быть использованы для нового синтеза белков и других соединений (пуриновых и пиримидиновых оснований, нуклеотидов, порфиринов и т.д.), для получения энергии (например, посредством включения в цикл трикарбоновых кислот) или могут быть подвергнуты дальнейшей деградации с образованием конечных продуктов А. о., подлежащих выведению из организма.
Аминокислоты, поступающие в составе белков пищи, используются для синтеза белков органов и тканей организма. Они участвуют также в образовании многих других важных биологических соединений: пуриновых нуклеотидов (глутамин, глицин, аспарагиновая кислота) и пиримидиновых нуклеотидов (глутамин, аспарагиновая кислота), серотонина (триптофан), меланина (фенилалпнин, тирозин), гистамина (гистидин), адреналина, норадреналина, тирамина (тирозин), полиаминов (аргинин, метионин), холина (метионин), порфиринов (глицин), креатина (глицин, аргинин, метионин), коферментов, сахаров и полисахаридов, липидов и т.д. Важнейшей для организма химической реакцией, в которой участвуют практически все аминокислоты, является трансаминирование, заключающееся в обратимом ферментативном переносе a -аминогруппы аминокислот на a -углеродный атом кетокислот или альдегидов. Трансаминирование является принципиальной реакцией биосинтеза заменимых аминокислот в организме. Активность ферментов, катализирующих реакции трансаминирования, - аминотрансфераз - имеет большое клинико-диагностическое значение.
Деградация аминокислот может протекать по нескольким различным путям. Большинство аминокислот способно подвергаться декарбоксилированию при участии ферментов декарбоксилаз с образованием первичных аминов, которые затем могут окисляться в реакциях, катализируемых моноаминоксидазой или диаминоксидазой. При окислении биогенных аминов (гистамина, серотонина, тирамина, g -аминомасляной кислоты) оксидазами образуются альдегиды, подвергающиеся дальнейшим превращениям, и аммиак , основным путем дальнейшего метаболизма которого является образование мочевины.
Другим принципиальным путем деградации аминокислот является окислительное дезаминирование с образованием аммиака и кетокислот. Прямое дезаминирование L-аминокислот в организме животных и человека протекает крайне медленно, за исключением глутаминовой кислоты, которая интенсивно дезаминируется при участии специфического фермента глутаматдегидрогеназы. Предварительное трансаминирование почти всех a -аминокислот и дальнейшее дезаминирование образовавшейся глутаминовой кислоты на a -кетоглутаровую кислоту и аммиак является основным механизмом дезаминирования природных аминокислот.
Продуктом разных путей деградации аминокислот является аммиак, который может образовываться и в результате метаболизма других азотсодержащих соединений (например, при дезаминировании аденина, входящего в состав никотинамидадениндинуклеотида - НАД). Основным путем связывания и нейтрализации токсичного аммиака у уреотелических животных (животные, у которых конечным продуктом А. о, является мочевина) служит так называемый цикл мочевины (синоним: орнитиновый цикл, цикл Кребса - Гензелейта), протекающий в печени. Он представляет собой циклическую последовательность ферментативных реакций, в результате которой из молекулы аммиака или амидного азота глутамина, аминогруппы аспарагановой кислоты и диоксида углерода осуществляется синтез мочевины. При ежедневном потреблении 100 г белка суточное выведение мочевины из организма составляет около 30 г . У человека и высших животных существует еще один путь нейтрализации аммиака - синтез амидов дикарбоновых кислот аспарагана и глутамина из соответствующих аминокислот. У урикотелических животных (рептилии, птицы) конечным продуктом А. о. является мочевая кислота.
В результате расщепления нуклеиновых кислот и нуклеопротеинов в желудочно-кишечном тракте образуются нуклеотиды и нуклеозиды. Олиго- и моно-нуклеотиды при участии различных ферментов (эстераз, нуклеотидаз, нуклеозидаз, фосфорилаз) превращаются затем в свободные пуриновые и пиримидиновые основания.
Дальнейший путь деградации пуриновых оснований аденина и гуанина состоит в их гидролитическом дезаминировании под влиянием ферментов аденазы и гуаназы с образованием соответственно гипоксантина (6-оксипурина) и ксантина (2,6-диоксипурина), которые затем превращаются в мочевую кислоту в реакциях, катализируемых ксантиноксидазой. Мочевая кислота - один из конечных продуктов А. о. и конечный продукт обмена пуринов у человека - выводится из организма с мочой. У большинства млекопитающих имеется фермент уриказа, который катализирует превращение мочевой кислоты в экскретируемый аллантоин.
Деградация пиримидиновых оснований (урацила, тимина) состоит в их восстановлении с образованием дигидропроизводных и последующем гидролизе, в результате которого из урацила образуется b -уреидопропионовая кислота, а из нее - аммиак, диоксид углерода и b -аланин, а из тимина - b -аминоизомасляная кислота, диоксид углерода и аммиак. Диоксид углерода и аммиак могут далее включаться в мочевину через цикл мочевины, а b -аланин участвует в синтезе важнейших биологически активных соединений - гистидинсодержащих дипептидов карнозина (b -аланил-L-гистидина) и анзерина (b -аланил-N-метил-L-гистидина), обнаруживаемых в составе экстрактивных веществ скелетных мышц, а также в синтезе пантотеновой кислоты и кофермента А.
Т.о., разнообразные превращения важнейших азотистых соединений организма связаны между собой в единый обмен. Сложный процесс А. о. регулируется на молекулярном, клеточном и тканевом уровнях. Регуляция А. о. в целом организме направлена на приспособление интенсивности А. о. к изменяющимся условиям окружающей и внутренней среды и осуществляется нервной системой как непосредственно, так и путем воздействия на железы внутренней секреции.
У здоровых взрослых людей содержание азотистых соединений в органах, тканях, биологических жидкостях находится на относительно постоянном уровне. Избыток азота, поступившего с пищей, выводится с мочой и калом, а при недостатке азота в пище нужды организма в нем могут покрываться за счет использования азотистых соединений тканей тела. При этом состав мочи изменяется в зависимости от особенностей А. о. и состояния азотистого баланса. В норме при неизменном режиме питания и относительно стабильных условиях окружающей среды из организма выделяется постоянное количество конечных продуктов А. о., а развитие патологических состояний приводит к его резкому изменению. Значительные изменения экскреции азотистых соединений с мочой, в первую очередь экскреции мочевины, могут наблюдаться и при отсутствии патологии в случае существенного изменения режима питания (например, при изменении количества потребляемого белка), причем концентрация остаточного азота (см. Азот остаточный ) в крови меняется незначительно.
При исследовании А. о. необходимо учитывать количественный и качественный состав принимаемой пищи, количественный и качественный состав азотистых соединений, выделяемых с мочой и калом и содержащихся в крови. Для исследования А. о. применяют азотистые вещества, меченные радионуклидами азота, фосфора, углерода, серы, водорода, кислорода, и наблюдают за миграцией метки и включением ее в состав конечных продуктов А. о. Широко используют меченые аминокислоты, например 15 N-глицин, которые вводят в организм с пищей или непосредственно в кровь. Значительная часть меченого азота глицина пищи выводится в составе мочевины с мочой, а другая часть метки попадает в тканевые белки и выводится из организма крайне медленно. Проведение исследования А. о. необходимо для диагностики многих патологических состояний и контроля за эффективностью лечения, а также при разработке рациональных схем питания, в т.ч. лечебного (см. Питание лечебное ).
Патологию А. о. (вплоть до очень значительной) вызывает белковая . Ее причиной может стать общее недоедание, продолжительный дефицит белка или незаменимых аминокислот в рационе, недостаток углеводов и жиров, обеспечивающих энергией процессы биосинтеза белка в организме. Белковая может быть обусловлена преобладанием процессов распада белков над их синтезом не только в результате алиментарного дефицита белка и других важнейших пищевых веществ, но и при тяжелой мышечной работе, травмах, воспалительных и дистрофических процессах, ишемии, инфекции, обширных ах, дефекте трофической функции нервной системы, недостаточности гормонов анаболического действия (гормона роста, половых гормонов, инсулина), избыточном синтезе или избыточном поступлении извне стероидных гормонов и т.п. Нарушение усвоения белка при патологии желудочно-кишечного тракта (ускоренная эвакуация пищи из желудка, гипо- и анацидные состояния, закупорка выводного протока поджелудочной железы, ослабление секреторной функции и усиление моторики тонкой кишки при энтеритах и энтероколитах, нарушение процесса всасывания в тонкой кишке и др.) также может приводить к белковой недостаточности. Белковая ведет к дискоординации А. о. и характеризуется резко выраженным отрицательным азотистым балансом.
Известны случаи нарушения синтеза определенных белков (см. Иммунопатология , Ферментопатии ), а также генетически обусловленного синтеза аномальных белков, например при гемоглобинопатиях , миеломной болезни (см. Парапротеинемические гемобластозы ) и др.
Патология А. о., заключающаяся в нарушении обмена аминокислот, часто связана с аномалиями процесса трансаминирования: уменьшением активности аминотрансфераз при гипо- или авитаминозах В 6 , нарушением синтеза этих ферментов, недостатком кетокислот для трансаминирования в связи с угнетением цикла трикарбоновых кислот при гипоксии и сахарном е и т.д. Снижение интенсивности трансаминирования приводит к угнетению дезаминирования глутаминовой кислоты, а оно, в свою очередь, - к повышению доли азота аминокислот в составе остаточного азота крови (гипераминоацидемии), общей гиперазотемии и аминоацидурии. Гипераминоацидемия, аминоацидурия и общая азотемия характерны для многих видов патологии А. о. При обширных поражениях печени и других состояниях, связанных с массивным распадом белка в организме, нарушаются процессы дезаминирования аминокислот и образования мочевины таким образом, что возрастают концентрация остаточного азота и содержание в нем азота аминокислот на фоне снижения относительного содержания в остаточном азоте азота мочевины (так называемая продукционная азотемия).
Продукционная азотемия, как правило, сопровождается выведением избытка аминокислот с мочой, поскольку даже в случае нормального функционирования почек фильтрация аминокислот в почечных клубочках происходит интенсивнее, чем их реабсорбция в канальцах. Заболевания почек, обтурация мочевых путей, нарушение почечного кровообращения приводят к развитию ретенционной азотемии, сопровождающейся нарастанием концентрации остаточного азота в крови за счет повышения содержания в крови мочевины (см. Почечная недостаточность ). Обширные раны, тяжелые и, инфекции, повреждения трубчатых костей, спинного и головного мозга, болезнь Иценко - Кушинга и многие другие тяжелые заболевания сопровождаются аминоацидурией. Она характерна и для патологических состояний, протекающих с нарушением процессов реабсорбции в почечных канальцах: болезни Вильсона - Коновалова (см. Гепатоцеребральная дистрофия ), нефронофтизе Фанкони (см. Рахитоподобные болезни ) и др. Эти болезни относятся к многочисленным генетически обусловленным нарушениям А. о. Избирательное нарушение реабсорбции цистина и цистинурия с генерализованным нарушением обмена цистина на фоне общей аминоацидурии сопровождает так называемый цистиноз. При этом заболевании кристаллы цистина откладываются в клетках ретикулоэндотелиальной системы. Наследственное заболевание фенилкетонурия характеризуется нарушением превращения фенилаланина в тирозин в результате генетически обусловленной недостаточности фермента фенилаланин - 4-гидроксилазы, что вызывает накопление в крови и моче непревращенного фенилаланина и продуктов его обмена - фенилпировиноградной и фенилуксусной кислот. Нарушение превращений этих соединений характерно и для вирусного гепатита.Тирозинемию, тирозинурию и тирозиноз отмечают при ах, диффузных заболеваниях соединительной ткани (коллагенозах) и других патологических состояниях. Они развиваются вследствие нарушения трансаминирования тирозина. Врожденная аномалия окислительных превращений тирозина лежит в основе алкаптонурии, при которой в моче накапливается непревращенный метаболит этой аминокислоты - гомогентизиновая кислота. Нарушения пигментного обмена при гипокортицизме (см. Надпочечники ) связаны с угнетением превращения тирозина в меланин вследствие ингибирования фермента тирозиназы (полное выпадение синтеза этого пигмента характерно для врожденной аномалии пигментации - а).
При массивном распаде клеточных структур (голодание, тяжелая мышечная работа, инфекции и др.) отмечают патологическое нарастание концентрации остаточного азота за счет увеличения относительного содержания в ней азота мочевой кислоты (в норме концентрация мочевой кислоты в крови не превышает - 0,4 ммоль/л ).
В пожилом возрасте снижаются интенсивность и объем синтеза белка за счет непосредственного угнетения биосинтетической функции организма и ослабления его способности усваивать аминокислоты пищи; развивается отрицательный азотистый баланс. Нарушения обмена пуринов у людей пожилого возраста приводят к накоплению и отложению в мышцах, суставах и хрящах солей мочевой кислоты - уратов. Коррекция нарушений А. о. в пожилом возрасте может быть осуществлена за счет специальных диет, содержащих полноценные животные белки, витамины и микроэлементы, с ограниченным содержанием пуринов.
Азотистый обмен у детей отличается рядом особенностей, в частности положительным азотистым балансом как необходимым условием роста. Интенсивность процессов А. о. на протяжении роста ребенка подвергается изменениям, особенно ярко выраженным у новорожденных и детей раннего возраста. В течение первых 3-х дней жизни азотистый баланс отрицателен, что объясняется недостаточным поступлением белка с пищей. В этот период обнаруживается транзиторное повышение концентрации остаточного азота в крови (так называемая физиологическая азотемия), иногда достигающее 70 ммоль/л ; к концу 2-й нед.
жизни концентрация остаточного азота снижается до уровня, отмечаемого у взрослых. Количество выделяемого почками азота нарастает в течение первых 3-х дней жизни, после чего снижается и вновь начинает увеличиваться со 2-й нед. жизни параллельно возрастающему количеству пищи.Наиболее высокая усвояемость азота в организме ребенка наблюдается у детей первых месяцев жизни. Азотистый баланс заметно приближается к равновесию в первые 3-6 мес. жизни, хотя и остается положительным. Интенсивность белкового обмена у детей достаточно высока - у детей 1-го года жизни обновляется около 0,9 г белка на 1 кг массы тела в сутки, в 1-3 года - 0,8 г/кг/ сут., у детей дошкольного и школьного возраста - 0,7 г/кг/ сут.
Средние величины потребности в незаменимых аминокислотах, по данным ФАО ВОЗ (1985), у детей в 6 раз больше, чем у взрослых (незаменимой аминокислотой для детей в возрасте до 3 мес. является цистин, а до 5 лет - и гистидин). Более активно, чем у взрослых, протекают у детей процессы трансаминирования аминокислот. Однако в первые дни жизни у новорожденных из-за относительно низкой активности некоторых ферментов отмечаются гипераминоацидемия и физиологическая аминоацидурия в результате функциональной незрелости почек. У недоношенных, кроме того, имеет место аминоацидурия перегрузочного типа, т.к. содержание свободных аминокислот в плазме их крови выше, чем у доношенных детей. На первой неделе жизни азот аминокислот составляет 3-4% общего азота мочи (по некоторым данным, до 10%), и лишь к концу 1-го года жизни его относительное содержание снижается до 1%. У детей 1-го года жизни выведение аминокислот в расчете на 1 кг массы тела достигает величин выведения их у взрослого человека, экскреция азота аминокислот, достигающая у новорожденных 10 мг/кг массы тела, на 2-м году жизни редко превышает 2 мг/кг массы тела. В моче новорожденных повышено (по сравнению с мочой взрослого человека) содержание таурина, треонина, серина, глицина, аланина, цистина, лейцина, тирозина, фенилаланина и лизина. В первые месяцы жизни в моче ребенка обнаруживаются также этаноламин и гомоцитруллин. В моче детей 1-го года жизни преобладают аминокислоты пролин и [гидр]оксипролин.
Исследования важнейших азотистых компонентов мочи у детей показали, что соотношение мочевой кислоты, мочевины и аммиака в процессе роста существенно изменяется. Так, первые 3 мес. жизни характеризуются наименьшим содержанием в моче мочевины (в 2-3 раза меньше, чем у взрослых) и наибольшей экскрецией мочевой кислоты. Дети в первые три месяца жизни выделяют 28,3 мг/кг массы тела мочевой кислоты, а взрослые - 8,7 мг/кг . Относительно высокая экскреция у детей первых месяцев жизни мочевой кислоты способствует иногда развитию мочекислого инфаркта почек. Количество мочевины в моче нарастает у детей в возрасте от 3 до 6 месяцев, а содержание мочевой кислоты в это время снижается. Содержание аммиака в моче детей в первые дни жизни невелико, но затем резко возрастает и держится на высоком уровне на протяжении всего 1-го года жизни.
Характерной особенностью А. о. у детей является физиологическая креатинурия. Креатин обнаруживается еще в амниотической жидкости; в моче он определяется в количествах, превышающих содержание креатина в моче взрослых, начиная с периода новорожденности и до периода полового созревания. Суточная экскреция креатинина (дегидроксилированного креатина) с возрастом увеличивается, в то же время по мере нарастания массы тела ребенка относительное содержание азота креатинина мочи снижается. Количество креатинина, выводимого с мочой за сутки, у доношенных новорожденных составляет 10-13 мг/кг , у недоношенных 3 мг/кг , у взрослых не превышает 30 мг/кг.
При выявлении в семье врожденного нарушения А. о. необходимо проведение медико-генетического консультирования .
Библиогр.: Березов Т.Т. и Коровкин Б.Ф. Биологическая химия, с. 431, М., 1982; Вельтищев Ю.Е. и др. Обмен веществ у детей, с. 53, М., 1983; Дудел Дж. и др. Физиология человека, пер. с англ., т. 1-4, М., 1985; Зилва Дж.Ф. и Пэннелл П.Р. Клиническая химия в диагностике и лечении, пер. с англ., с. 298, 398, М., 1988; Кон Р.М. и Рой К.С. Ранняя диагностика болезней обмена веществ, пер. с англ., с. 211, М., 1986; Лабораторные методы исследования в клинике, под ред. В.В. Меньшикова, с. 222, М., 1987; Ленинджер А. Основы биохимии, пер. с англ., т. 2, М., 1985; Мазурин А.В. и Воронцов И.М. Пропедевтика детских болезней, с. 322, М., 1985; Руководство по педиатрии, под. ред. У.Е. Бермана и В.К. Вогана, пер. с англ., кн. 2, с. 337, VI., 1987; Страйер Л. Биохимия, пер. с англ., т. 2, с. 233, М., 1985.
Речь пойдет об особенностях метаболизма пуриновых оснований. Большинству людей это ни о чем не говорит. Но если вам знакомы слова «подагра», мочекаменная болезнь, инсулинорезистентность, сахарный диабет 2 типа, то знать суть о метаболизме пуринов необходимо. Казалось бы: а хирургия то здесь причем? А притом, что многие специалисты при болях в суставах и высокой мочевой кислоте ставят диагноз «подагра». На самом деле — все намного сложнее. К примеру подагрический артрит может быть при нормальных цифрах мочевой кислоты, и наоборот: высокая мочевая кислота может быть в ряде случаев у здорового человека.
Организм человека в основном состоит из четырех химических элементов, на долю которых приходится 89 % состава: С-углерод (50%), О-кислород(20%), Н-водород(10%) и N-азот (8,5%). Далее идет ряд макроэлементов: кальций, фосфор, калий, сера, натрий, хлор и др. Затем микроэлементы, количество которых очень мало, но они жизненно необходимы: марганец, железо, йод и пр.
Интересен нам будет четвертый в этом количественном списке — азот.
Живой организм — это динамическая система. По простому: вещества в него постоянно поступают (становясь частью организма) и выводятся из него. Основной источник азота для организма — белки. Поступающий с пищей белок в желудочно-кишечном тракте распадается до аминокислот, которые уже и включаются в обмен. Ну а каким образом азотсодержащие вещества выводятся из организма?
В процессе эволюции у животных выработались определенные особенности азотистого обмена.
Причем ключевым в определении этих особенностей будут: условия существования и доступ к воде.
Животных разделяют на три группы, имеющие различия в метаболизме азота:
Аммонио-литические
. Конечный продукт азотистого обмена — аммиак, NH3. Сюда относят большую часть водных беспозвоночных и рыб.
Все дело в том, что аммиак — токсичное вещество. И для его выведения нужно очень-очень много жидкости. Благо — он хорошо растворим в воде. С выходом на сушу в ходе эволюции возникла потребность в изменении метаболизма. Так появились:
Уреолитические . У этих животных появился так называемый «цикл мочевины». Аммиак связывается с СО2(углекислый газ). Образуется конечный продукт — мочевина. Мочевина не такое токсичное вещество и для ее выведения требуется заметно меньше жидкости. Кстати мы с вами относимся именно к этой группе. Мочевая кислота в процессе метаболизма в значительно меньших количествах также образуется, но распадается до малотоксичного и хорошо растворимого аллантоина. Но… Кроме человека и человекообразных обезьян. Это очень важно и к этому вернемся.
Урикотелические
. Предкам земноводных с уреолитическим обменом пришлось приспосабливаться к засушливым регионам. Это пресмыкающиеся и прямые предки динозавров — птицы. У них конечным продуктом является мочевая кислота. Она очень плохо растворяется в воде и для ее выведения из организма как раз воды много и не требуется. В помете у тех же птиц количество мочевой кислоты очень большое, фактически выводится в полутвердом виде Поэтому птичий помет («гуано») — основная причина коррозии и разрушения металлоконструкций мостов. Лакокрасочное покрытие автомобиля тоже портит — будьте внимательны, мойте сразу.
Это классическая гексагональная долька печени. В общем так печень выглядит под микроскопом. Похожа на Москву-сити, только вместо кремля — центральная вена. А интересовать нас будут «домики», плотно прилежащие друг к другу. Это гепатоциты — ключевые клетки печени.
Славянское слово печень произошло от слова «печь». Действительно, температура органа на градус выше температуры тела. Причина в этом — очень активный обмен веществ в гепатоцитах. Клетки действительно уникальные, в них протекает около 2 тысяч химических реакций.
Печень — это основной орган, который продуцирует мочевую кислоту. 95% выводимого азота — синтез мочевой кислоты как конечный продукт химических реакций в печени
. И только 5% — окисление пуриновых оснований, поступающих извне с пищей. Поэтому коррекция питания при гиперурикемии не является ключевым в лечении.
Схема обмена мочевой кислоты
 Откуда берутся пурины?
Откуда берутся пурины?
1. Пурины, которые поступают с пищей
. Как уже отмечалось — это небольшое количество — около 5%. Те пурины, которые содержаться в пище (больше всего, разумеется в печени и почках, красном мясе).
2. Синтез пуриновых оснований самим организмом
. Большая часть синтезируется в гепатоцитах печени. Очень важный пункт, к нему вернемся. А также причем здесь рекомендуемая диабетиками и не требующая для усвоения инсулина фруктоза.
3. Пуриновые основания, которые образуются в организме вследствие распада тканей: при онкопроцессах, псориазе
. Почему у спортсменов может повышаться мочевая кислота? Это и есть третий путь. Тяжелые физические нагрузки приводят к усилению процессов распада и синтеза тканей. Если вы накануне занимались тяжелым физическим трудом, а утром вы сдаете анализ, уровень мочевой кислоты может быть выше вашего среднего значения. Знакомимся: аденин и гуанин. Это и есть пуриновые основания. Совместно с тимином и цитозином формируют спираль ДНК. Студенты медики не любят — зубрежка на курсе биохимии:). Как известно, ДНК состоит из двух цепочек. Напротив аденина всегда становится тимин, напротив гуанина — цитозин. Две цепочки ДНК склеиваются как две половинки застежки-молнии. Количество этих веществ повышается при активном распаде тканей, как бывает, например, при онкопроцессах
Знакомимся: аденин и гуанин. Это и есть пуриновые основания. Совместно с тимином и цитозином формируют спираль ДНК. Студенты медики не любят — зубрежка на курсе биохимии:). Как известно, ДНК состоит из двух цепочек. Напротив аденина всегда становится тимин, напротив гуанина — цитозин. Две цепочки ДНК склеиваются как две половинки застежки-молнии. Количество этих веществ повышается при активном распаде тканей, как бывает, например, при онкопроцессах
Рядом последовательных химических реакций пурины преобразуются в мочевую кислоту.

Метаболизм мочевой кислоты у человека и приматов
Планировал максимально упростить для понимания схему. Пусть учат студенты-медики на 2 курсе:). Но названия ферментов оставил. Самый важный момент — фермент ксантиноксидаза
. Именно его активность падает при лечении аллопуринолом
(точнее эффективность, так как аллопуринол с ним конкурирует за рецептор), чем и снижается синтез мочевой кислоты.
Редко, но всречаются врожденное заболевание,сопровождающееся генетическим нарушением в синтезе ксантиноксидазы, при котором уровень мочевой кислоты снижен. В таком случае накапливаются ксантин и гипоксантин. Ксантинурия. Казалось бы ну и хорошо, меньше мочевой кислоты. Однако выяснилось, что мочевая кислота не только вредна, но и полезна…
Разговор о вреде и пользе мочевой кислоты следует начать очень издалека. Тогда, 17 миллионов лет назад, в эпоху миоцена у наших предков произошла мутация в гене, который продуцирует фермент — уриказу. И нам досталась «урезанная» версия пуринового обмена.
 У других млекопитающих уриказа переводит мочевую кислоту в растворимый и легко выводящийся из организма аллантоин. И у этих животных никогда не бывает подагры. Может возникнуть предположение, что в этой мутации нет никакого смысла. Но эволюция этот ген не исключила: мутация оказалась необходимой.
У других млекопитающих уриказа переводит мочевую кислоту в растворимый и легко выводящийся из организма аллантоин. И у этих животных никогда не бывает подагры. Может возникнуть предположение, что в этой мутации нет никакого смысла. Но эволюция этот ген не исключила: мутация оказалась необходимой.
Современные исследования показали, что мочевая кислота является побочным продуктом разложения фруктозы в печени и накопление солей мочевой кислоты способствует эффективному превращению фруктозы в жир. Таким образом, у наших предков в геноме закрепился ген «бережливости». Тогда ген был необходим для создания запасов на голодный период. Было доказано, что окончательная инактивация уриказы совпала с глобальным похолоданием климата на Земле. Нужно было «наесть» как можно больше запасов подкожного жира на холодный период, перевести содержащуюся в плодах фруктозу в жировой запас. Сейчас проводятся ряд экспериментов с введением в клетки печени фермента уриказы. Не исключено, что в дальнейшем на основе фермента уриказы появятся препараты для лечения подагры. Так что склонность к ожирению у нас заложена в генах. На несчастье тем многим мужчинам и женщинам, страдающим полнотой. Но проблема не только в генетике. Изменился характер питания современного человека.
Про вред и пользу мочевой кислоты, а также про питание при гиперурикемии
Известно, что постоянный уровень мочевой кислоты способен значительно повысить риск ряда заболеваний. Однако доказано, что периодическое повышение уровня мочевой кислоты может оказывать положительное действие. Исторически доступ к мясной пище (основному источнику пуринов), был нерегулярным. Основная пища: различные коренья, плоды деревьев. Ну а если принесет первобытный охотник добычу — так это праздник. Поэтому, периодическое от мясных продуктов было обычным образом жизни. Есть добыча — едим до отвала. Нет добычи — едим растительную пищу. Сейчса установлено, что кратковременное, периодическое повышение уровня мочевой кислоты благоприятно вляет на развитие и функцию нервной системы. Может поэтому и начал развиваться мозг?
Как эта мочевая кислота выводится из организма
Пути два: почки и печень
Основной путь — выведение с почками — это 75%
25 процентов выводится печенью с помощью желчи. Поступившая в просвет кишечника мочевая кислота и разрушается (спасибо нашим бактериям в кишечнике).
В почки мочевая кислота попадает в виде натриевой соли. При ацидозе (закислении мочи) в почечных лоханках могут формироваться микролиты. Тот самый «песок» и «камни». Кстати алкоголь очень сильно снижает экскрецию уратов с мочой. Почему и приводит к приступу подагры.
Итак, какой нужно сделать вывод?Методы снижения мочевой кислоты
1. Стараться в неделю 1-2 дня делать чисто вегетарианским
2. Наибольшее количество пуринов содержится в тканях животного происхождения. Причем в животных клетках с активным метаболизмом: печени, почках — больше всего.
3. Нужно есть меньше жирной пищи, так как избыток насыщенных жиров подавляет способность организма перерабатывать мочевую кислоту.
4. Едим поменьше фруктозы. Мочевая кислота — продукт метаболизма фруктозы. Ранее пациентам с сахарным диабетом рекомендовали заменять глюкозу на фруктозу. Действительно, фруктоза для своего усвоения не требует участия инсулина. Но для усвоения фруктоза еще тяжелее. Внимание: в сахаре молекула сахарозы — это дисахарид — глюкоза + фруктоза. Так что сахара едим меньше.
5. Исключить прием алкоголя, особенно пива. Вино в небольших количествах не влияет на уровень мочевой кислоты.
6. Очень интенсивные физические нагрузки повышают уровень мочевой кислоты.
7. Нужно пить много воды. Это позволит эффективно выводить мочевую кислоту.
Если у вас повышена мочевая кислота
Ну во первых, к счастью это не всегда является патологией: кратковременный подъем может быть вариантом нормы
Если все же проблема есть, нужно разобраться, на каком уровне есть нарушение (та самая первая схема): нарушения в синтезе пуринов (тот самый метаболический синдром), алиментарный фактор (много мяса кушаем, пивом запиваем), нарушение функции почек (нарушение экскреции мочевой кислоты)или сопутствующие заболевания, сопровождающиеся разрушением тканей.
Удачи Вам и грамотных докторов.
Если вы нашли опечатку в тексте, пожалуйста, сообщите мне об этом. Выделите фрагмент текста и нажмите Ctrl+Enter .
